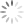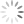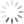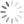 (Carregando Índice)... (Carregando Índice)... |
Você está em:
Inicial  acp-medicine Imunologia
acp-medicine Imunologia
Última revisão: 12/05/2016
Comentários de assinantes: 0
Steven K. Lundy, PhD, David A. Fox, MD e Alison Gizinski, MD
Artigo original: Lundy SK, PhD, Fox DA, MD, Gizinski A, MD. Introduction to clinical immunology: overview of the immune response, autoimmune conditions, and immunosuppressive therapeutics. ACP Medicine. 2015.
[The original English language work has been published by DECKER INTELLECTUAL PROPERTIES INC. Copyright © 2015 Decker Intellectual Properties Inc. All Rights Reserved.]
Tradução: Paulo Henrique Machado.
Revisão técnica: Dr. Lucas Santos Zambon
O sistema imune é uma rede complexa de células e mediadores cuja finalidade é manter em equilíbrio a tarefa de proteger hospedeiros contra ameaças invasivas, incluindo microrganismos patogênicos, parasitas, células transformadas e toxinas, mantendo a tolerância aos autotecidos e dos micróbios comensais. A ampla diversidade de microrganismos aos quais os seres humanos estão expostos, assim como sua rápida capacidade de mutação e de subverter as respostas imunes, aumenta a necessidade de um sistema imune altamente dinâmico e adaptável com uma sofisticada rede de comunicação. Mesmo assim, muitos micróbios encontrados na vida cotidiana não são patogênicos devido à baixa capacidade para invadir ou para infectar os hospedeiros, e para o funcionamento apropriado do sistema imune para limitar sua patogenicidade. As respostas imunes ativas podem ser tóxicas para os tecidos do corpo humano e exigem muita energia e nutrientes para expandir as populações de células efetoras e sintetizar os mediadores imunes. Esses fatores criam pressões seletivas sobre o sistema imune para gerar respostas somente quando os indivíduos forem colocados em situação de perigo, para particularizar a reação e melhorar o confronto e neutralizar o agente causativo, e para interromper a atividade a partir da eliminação do perigo. Sistemas imunes com funcionamento adequado protegem a homeostase por meio da sensibilidade de quando e como responder a situações ameaçadoras e através do uso de sistemas sofisticados de pontos de verificação.
Sob a perspectiva clínica, muitas doenças e condições mantêm uma ligação óbvia com o funcionamento inadequado do sistema imune. Respostas imunes insuficientes poderão produzir infecções agudas e crônicas incontroláveis. Entretanto, respostas manifestas do sistema imune a infecções, como aquelas observadas nos casos de choque séptico ou da síndrome do desconforto respiratório agudo (SDRA), também podem ter consequências fatais. Provavelmente, as respostas desnecessárias a materiais estranhos inócuos ou aos autoantígenos sejam os deflagradores subjacentes de alergias e autoimunidade, respectivamente. Além do papel clássico desempenhado nessas doenças infecciosas e inflamatórias, o sistema imune tem também uma participação importante na vigilância e no controle de tumores, doenças cardiovasculares, complicações da saúde relacionadas à obesidade, doenças neuromusculares, depressão e algumas formas de demência. Consequentemente, o conhecimento do papel desempenhado pela imunidade em processos de doença está se tornando cada vez mais importante na maior parte dos aspectos ou mesmo em todos os aspectos da prática clinica. Neste capítulo, apresentaremos os componentes especiais do sistema imune tendo em vista que se relacionam a tipos específicos de respostas imunes e ao contexto da patogênese da doença e, em seguida, apresentaremos uma discussão sobre os alvos do tratamento clínico e uma visão geral dos agentes terapêuticos utilizados no controle da autoimunidade.
|
As informações financeiras estão no final deste capitulo, antes das referências. |
As reações imunes normais podem ser desmembradas em quatro etapas principais. O primeiro passo é detectar a presença de algum antígeno estranho ou de um estímulo imune entre todos os estímulos potenciais com origem no hospedeiro (autoantígenos). Na maior parte dos casos, essa detecção é feita por meio de um instrumento especializado no reconhecimento de antígenos nas células do sistema imune e através de um sistema complexo de “educação” cuja função é eliminar ou suprimir o reconhecimento de autoantígenos. O segundo passo é determinar se a eventual presença de alguma fonte de antígenos estranhos está ameaçando o hospedeiro. Essa etapa é extremamente importante porque os seres humanos se expõem a muitos microrganismos e a outros estímulos, em especial às bactérias comensais e aos fungos ou moléculas associadas aos alimentos, para os quais a reação imune talvez seja inadequada e potencialmente prejudicial. Com frequência, determina-se o nível de periculosidade de um estímulo de acordo com a localização e o contexto. Portanto, uma bactéria comensal na pele possivelmente seja considerada segura e é ignorada pelo sistema imune, porém ela deverá ser detectada como uma ameaça potencial nos casos em que penetrar na corrente sanguínea. A ativação do grupo correto de células e dos mediadores imunes para tratar o estímulo de uma forma rápida e eficiente é o terceiro passo para a obtenção de respostas imunes. A resposta pró-inflamatória é um dispositivo deflagrador precoce e fundamental que dispara a ativação de respostas imunes. Em seguida, o sistema imune interpreta e integra diversos sinais para calcular qual entre suas várias ramificações e componentes precisa ser envolvido no processo para controlar o agente desencadeador. Os componentes celulares e moleculares que formam essas ramificações do sistema imune serão apresentados com mais detalhes mais adiante. A coordenação efetiva desses componentes é essencial para o sucesso e a eficiência da proteção de hospedeiros. A etapa final em respostas imunes normais é o retorno para um estado homeostático logo após a eliminação ou controle de uma ameaça. Esse retorno ocorre através de mecanismos passivos relacionados à ausência do agente desencadeador e por meio da supressão imune ativa mediada por células e mediadores especiais que se localizam nas ramificações reguladoras do sistema imune.
A maior parte das células do sistema imune se desenvolve a partir de células-tronco hematopoiéticas (CTHs) pluripotentes que residem na medula óssea. A diferenciação inicial das CTHs resulta na produção das populações de células do progenitor linfoide comum (PLC) ou do progenitor mieloide comum (PMC). A seguir, os PLCs diferenciam nos três tipos principais de linfócitos: linfócitos T, linfócitos B e células destruidoras naturais (KCs, do inglês killer cells), ou conhecidas como “natural killers” (NK). A diferenciação dos PMCs forma células dendríticas imaturas, progenitores de granulócitos/monócitos ou linhagens de progenitores de megacariócitos/eritrócitos. As plaquetas e os eritrócitos são produtos de megacariócitos e eritroblastos, que são produtos derivados dos progenitores de megacariócitos/eritrócitos. As populações principais de células imunes granulocitárias - leucócitos polimorfonucleares (PMNs, do inglês polymorphonuclear neutrophilic leukocytes), eosinófilos, basófilos e mastócitos – derivam de progenitores de granulócitos/monócitos. Como alternativa, os progenitores de granulócitos/monócitos podem produzir monócitos que circulam na corrente sanguínea e, mais tarde, se tornam terminalmente diferenciados em macrófagos no momento em que atingem os tecidos periféricos. A Tabela 1 apresenta um resumo da classificação e das principais características funcionais desses tipos de células com origem na medula óssea.
As exceções de células imunes com origem na medula óssea são subgrupos de células imunes que estão presentes no nascimento, muitas das quais são produzidas a partir das células-tronco encontradas no fígado fetal.1 Essas células imunes pré-natais e os respectivos fatores associados, que presumivelmente surgem sem estímulos antigênicos externos, fazem parte de um ramo maior do sistema imune que faz a mediação da imunidade inata. Em termos mais gerais, imunidade inata se refere às respostas imunes com foco amplo nas classes principais de microrganismos, ao invés de se direcionarem especificamente a um patógeno ou antígeno individual.
|
Tabela 1: Classificação e Principais Características Funcionais das Células dos Sistema Imune
| |||
|
Categoria |
População de Células Efetoras
|
Ramo Funcional |
Funções Efetoras Principais |
|
Células T |
Linfócito Th1 |
Adaptativo |
Respostas imunes diretas aos micróbios intracelulares; segrega interferon gama, TNT-a, IL-2.
|
|
|
Linfócito Th2 |
Adaptativo |
Respostas imunes diretas aos micróbios extracelulares; segrega IL-4, IL-5, IL-13.
|
|
|
Linfócito Th17 |
Adaptativo |
Respostas diretas na mucosa contra os micróbios extracelulares; segrega IL-17, IL-22.
|
|
|
Célula T reguladora natural (Treg) |
Adaptativo |
Suprime a autoimunidade e outras reações inflamatórias; segrega IL-10, TGF-ß.
|
|
|
Treg induzida |
Adaptativo |
Suprime inflamações depois de infecções; segrega IL-10, TGF-ß.
|
|
|
Célula T auxiliar folicular (Tfh)
|
Adaptativo |
Maturação direta de anticorpos por meio da coestimulação de células B em centros germinais. |
|
|
Linfócitos T citotóxicos (LTC)
|
Adaptativo |
Mata vírus e células infectadas por bactérias intracelulares; vigilância de tumores. |
|
|
Células T NK invariantes
|
Inato |
Produção precoce de citocina de células T em resposta aos antígenos glicolipídeos. |
|
APCs |
Células dendríticas |
Inato e indutor de adaptativo
|
Apresentam antígenos e coestimulam células Th virgens; segregam citocinas inatas. |
|
|
Linfócito B-1 |
Inato e indutor de adaptativo
|
Produz anticorpos naturais; apresenta antígenos para as células Th. |
|
|
Linfócito B-2 |
Adaptativo
|
Produz a maior parte dos anticorpos em circulação; apresenta antígenos para as células Th.
|
|
|
Monócito/macrófago |
Inato e indutor de adaptativo
|
Fagocitose, apresentação de antígenos, secreção de citocinas inatas. |
|
Granulócitos |
Basófilo |
Inato |
Liberação granular na direção de micróbios extracelulares; secreção de IL-4.
|
|
Tabela 1: Classificação e Principais Características Funcionais das Células dos Sistema Imune
| |||
|
Categoria |
População de Células Efetoras
|
Ramo Funcional |
Funções Efetoras Principais |
|
|
Mastócito |
Inato |
Proteção da pele e da mucosa contra micróbios extracelulares.
|
|
|
Eosinófilo |
Inato |
Liberação granular em resposta a vermes e parasitas; induz secreção de muco.
|
|
|
Neutrófilo |
Inato |
Eliminação de bactérias e de fungos por fagocitose.
|
|
Outros |
Célula NK |
Inato |
Mata vírus e células infectadas por bactérias intracelulares; vigilância de tumores.
|
IL = interleucina; NK – natural killer; PMN (polymorphonuclear neutrophilic leukocytes) = leucócitos polimorfonucleares; Th = T auxiliar; TGF (transforming growth factor) = fator de crescimento transformador; TNF (tumor necrosis factor) = fator de necrose tumoral.
As células de imunidade inata estão presentes o tempo todo e em todos os indivíduos, independente da exposição prévia aos micróbios aos quais reagem. Os mediadores e as células do sistema imune inato são pré-programados para responder aos sinais microbianos e permitir respostas imediatas. Os mediadores solúveis de imunidade inata incluem anticorpos “naturais” e componentes complementares. Os anticorpos naturais estão presentes no nascimento e têm amplas especificidades para as classes mais importantes de micróbios. As respostas imunes inatas não aumentam com exposições sucessivas a um determinado tipo de patógeno e, portanto, não são alvos específicos das vacinações. As células que geralmente são classificadas como parte do sistema imune inato incluem granulócitos, monócitos/macrófagos e células dendríticas. Além disso, diversos subgrupos menores da linhagem de linfócitos – linfócitos B1, células T gama/delta e células destruidoras naturais invariantes (iNKT) – são considerados partes integrantes do sistema imune inato, com base em suas características evolutivas exclusivas e nas amplas especificidades. As células e as moléculas do sistema imune inato participam também de respostas imunes adaptativas.
Talvez seja mais fácil compreender o termo imunidade inata em comparação com imunidade adaptativa, que se refere ao ramo do sistema imune que é ativado na direção de espécies individuais de microrganismos, antígenos ou toxinas somente depois de tê-los encontrado. Os linfócitos são os componentes celulares mais importantes do sistema imune adaptativo e se classificam com base na maturação no timo (linfócitos T) ou pela capacidade de produzir anticorpos solúveis altamente específicos (linfócitos B-2). Ao contrário da imunidade inata, o desenvolvimento das respostas imunes adaptativas é mais lento, porém, mesmo assim, conferem proteção duradoura e memória imunológica. A imunidade adaptativa envolve a ativação e a expansão de subgrupos de linfócitos que são direcionados contra as proteínas ou outras biomoléculas produzidas somente por tipos específicos de micróbios. Posteriormente, os linfócitos T se subdividem de acordo com os respectivos marcadores moleculares e com as funções efetoras em linfócitos T (Th) auxiliares (células Th) e linfócitos T citotóxicos (LTCs). As células se especializam na integração de sinais distintos produzidos durante tipos diferentes de infecções, direcionando as respostas imunes corretas das outras células para aqueles estímulos. Portanto, essas células são mediadoras das respostas imunes adaptativas e ocupam uma posição extremamente importante no sistema imune. Os mecanismos através dos quais as células do sistema imune adaptativo reconhecem os antígenos e seus mediadores solúveis serão discutidos com mais detalhes em outras seções deste capítulo.
É importante observar que a maioria das respostas imunes, dos mediadores e das células imunes inatas e adaptativas, opera de forma simultânea e cooperativa no combate aos patógenos. Como enfatizaremos nas seções seguintes, as células imunes adaptativas dependem das células do sistema imune inato no processo de apresentação de antígenos para iniciar sua ativação. Por outro lado, as células imunes adaptativas direcionam as respostas das células imunes inatas e, às vezes, armam essas células com moléculas efetoras, que podem conferir um nível elevado de especificidade antigênica. No centro dessa regulação cruzada entre imunidade inata e adaptativa estão os mecanismos de processamento e de reconhecimento e os mecanismos altamente complexos da comunicação intercelular.
Os princípios orientadores sobre como as células do sistema imune interagem entre si e com o ambiente estão relacionados com o reconhecimento do padrão molecular. Esse processo se refere à interação entre os receptores da superfície celular da célula que recebe o sinal e os ligantes específicos, que podem ser moléculas solúveis ou componentes da superfície celular. Usualmente, as interações entre receptores e ligantes no sistema imune se caracterizam pelos contatos entre proteínas; no entanto, lipídeos, carboidratos e ácidos nucleicos também podem ser reconhecidos por receptores do sistema imune. Em um determinado momento, alguma célula individual poderá coletar amostras da célula associada e dos ligantes solúveis que estiverem mais próximos para verificar a eventual presença de quaisquer sinais de ativação. A seguir, esses sinais são integrados e processados por séries complexas de moléculas sinalizadoras intracelulares que, em última análise, determinam se a célula irá modificar os padrões de expressão gênica, iniciar a divisão celular, atacar algum microrganismo, sofrer morte celular programada ou se tornar ou permanecer inativa.
Muitas células têm receptores especiais de reconhecimento de padrões (PRRs, do inglês pattern recognition receptors) que conseguem fazer o reconhecimento direto das estruturas geralmente expressas por microrganismos. Um exemplo típico de PRRs é a família de receptores toll-like (TLR, do inglês toll-like receptors), cujos ligantes incluem componentes das paredes de células bacterianas e fúngicas (TLRs 1, 2, 4 e 6), RNA viral de fita dupla (TLR3), RNA viral de fita única (TLR7 e TLR8) ou DNA bacteriano e viral não metilado (TLR9). De maneira geral, a ativação de PRRs é o primeiro sinal da penetração de algum micróbio potencialmente perigoso no hospedeiro, sendo que a ativação subsequente da célula que expressa o PRR dispara respostas inflamatórias. Outra família de PRRs forma um complexo conhecido por inflamassomo (ou inflamassoma) no interior do citoplasma da célula, sendo que tanto a família de TLRs como o inflamassomo utiliza o fator nuclear kB (NFkB, do inglês nuclear factor kappa B) do fator de transcrição como mediador principal dos respectivos efeitos pró-inflamatórios.3 Os PRRs são classificados como componentes da imunidade inata por causa das amplas especificidades, conferida por estruturas codificadas pelo DNA que está em sua configuração na linha geminal. Entretanto, os PRRs também são expressos por células do sistema imune adaptativo, principalmente os linfócitos B.
Ao contrário dos TLRs e de outros receptores expressos por células imunes inatas, a especificidade das células do sistema imune adaptativo é determinada por recombinações genéticas geradas aleatoriamente a partir dos genes receptores de antígenos. A maturação precoce do linfócito B-2 ocorre na medula óssea, sendo que, em parte, é estimulada por recombinações produtivas do lócus da imunoglobulina, resultando na expressão de uma molécula do anticorpo IgM na superfície celular. O anticorpo IgM superficial constitui o receptor de antígenos de células B (BCR, do inglês B cell receptors), sendo que, em parte, sua especificidade inicial depende da divisão diferencial de segmentos gênicos com a denotação V, D e T, com uma cadeia pesada µ de região constante, assim como deleções e inserções de nucleotídeos que ocorrem no momento da recombinação de genes e do desenvolvimento da medula óssea.4 Cada cadeia pesada forma um par com uma cadeia leve kappa ou lambda, que apresenta também uma diversidade inerente substancial. Essas alterações genéticas, que ocorrem em regiões determinantes da complementaridade (CDRs, do inglês complementary-determining regions) altamente variáveis, fazem com que cada linfócito B-2 tenha uma especificidade antigênica exclusiva com base na ligação direta de seu BCR com os ligantes expressos em outras superfícies celulares ou moléculas solúveis. O desenvolvimento da célula B-2 tem como suporte as células estromais da medula óssea, sendo que a seleção positiva de células B-2 com recombinações produtivas de uma molécula de BCR permite que as células B saiam da medula óssea e entrem em circulação.
Uma das características exclusivas da maturação das células B-2 é que poderão ocorrer recombinações genéticas ulteriores dentro do lócus de imunoglobulinas, resultando em alterações moleculares no anticorpo e, particularmente, em sua especificidade antigênica. Essas alterações são estimuladas por sinais de reconhecimento e de ativação de antígenos das células Th e pelo microambiente onde as respostas estiverem em curso. Os folículos de células B dos centros germinativos nos órgãos linfoides secundários (baço e linfonodos) são os sítios de maturação mais importantes das células B. Nesses sítios, as células B encontram os antígenos que permanecem sobre a superfície das células dendríticas foliculares especiais e, simultaneamente, recebem sinais de ativação e de proliferação das células auxiliares (Th, do inglês helper cells) foliculares residentes. Troca isotípica se refere à substituição de segmentos diferentes de uma região constante no lugar da cadeia µ na molécula de imunoglobulina, resultando na produção de formas solúveis alternativas do anticorpo (subclasses de IgG, IgE, IgA) que circulam afastando-se das células B totalmente diferenciadas que produzem anticorpos (células plasmáticas). A ligação da molécula CD40 da superfície celular por seu ligante nas células Th ativadas age como um desencadeador da troca isotípica, enquanto que o padrão de citocinas presente durante essa interação controla a produção de isotipos. A seção sobre os subgrupos de células Th apresenta informações mais detalhadas sobre padrões específicos de citocinas e de isotipos de anticorpos. As células B-2 ativadas também sofrem hipermutação somática do gene da imunoglobulina nas sequências da codificação da CDR, que poderá resultar em alterações produtivas ou improdutivas na especificidade da ligação do anticorpo. Nas células B, as alterações produtivas causadas por hipermutação somática são favorecidas, tendo em vista que possuem especificidade mais elevada do anticorpo para antígenos, resultando em um sinal mais potente para a proliferação celular no encontro com o antígeno e na consequente maturação da resposta imune na direção de um determinado tipo de patógeno.
Além do papel desempenhado no reconhecimento de antígenos, como a imunoglobulina superficial nos linfócitos B que os produzem, os anticorpos também são importantes na mediação de respostas das células que não expressam seus próprios anticorpos, incluindo as células associadas à imunidade inata. A região constante (Fc) que se localiza na cauda de isotipos solúveis de anticorpos (subclasses de IgG e IgE) pode se ligar por meio de receptores de Fc específicos para isotipos (FcRs) expressos na superfície de algumas células. Essa ligação aos anticorpos confere especificidade ao antígeno para a célula Fc+ e facilita algumas funções do efetor das células imunes inatas, incluindo a degranulação de granulócitos (mastócitos, basófilos e eosinófilos) e a citotoxicidade celular dependente de anticorpos por linfócitos destruidores naturais (células destruidoras naturais).
Os genes do receptor da célula T (TCR, do inglês T cell receptor) passam por recombinações genéticas que se assemelham àquelas descritas para os anticorpos. Combinações aleatórias de segmentos do gene VDJ resultam na formação de domínios de ligações antigênicas que conferem especificidade antigênica para cada célula T. Ao contrário dos anticorpos, os TCRs reconhecem antígenos somente quando se apresentarem no contexto molecular do complexo de histocompatibilidade principal (MHC) (também conhecido por antígeno leucocitário humano [HLA]). O desenvolvimento das células T ocorre no timo e é estimulado por células epiteliais tímicas que expressam MHCs em suas superfícies celulares. A seleção positiva de células T ocorre no córtex do timo, onde somente as células T com interação produtiva entre TCR e MHC recebem sinais de sobrevivência [ver a Figura 1]. As interações adicionais ocorrem no momento da seleção positiva entre o MHC de classe I ou classe II com as moléculas CD8 ou CD4, que são coexpressas através do desenvolvimento das células T, determinam se a célula T em fase de maturação irá assumir as características de um linfócito T citotóxico (LTC) CD8+ ou de um fenótipo do linfócito T auxiliar (célula Th) CD4+, respectivamente. Levando-se em consideração que os antígenos, para os quais muitas das células T selecionadas positivamente são específicas, não estão presentes no timo, esse processo depende principalmente das interações de receptor e ligante entre TCR e MHC. O LTC e as células T auxiliares que sobreviverem à seleção positiva poderão migrar para a medula tímica, onde passam por um processo de seleção negativa cuja função é proteger contra células autorreativas.
No processo de geração aleatória de receptores antigênicos produz-se um número significativo de linfócitos T e B com especificidades direcionadas para os autoantígenos normais dos indivíduos. Essas células autorreativas são uma grave ameaça para os indivíduos porque há uma grande probabilidade de que encontrem seus antígenos que, se forem ativados, produzem autoimunidade. Para proteger contra essa possibilidade, as células imunes adaptativas passam por diversos processos de seleção negativa durante seu desenvolvimento cuja finalidade é eliminar as células autorreativas ou forçá-las a assumir um papel regulador. Com frequência, as células T e B que reagirem fortemente contra os autoantígenos durante os processos evolutivos descritos acima sofrerão morte celular induzida por ativação (MCIA), resultando em tolerância imune central. Para as células T, o primeiro estágio de seleção negativa ocorre no momento em que houver uma interação muito intensa do TCR com os antígenos apresentados por células epiteliais tímicas ou por células medulares tímicas que expressam autoantígenos. A ativação resultante torna essas células sensíveis aos sinais indutores de morte presentes no timo. A importância da tolerância imune central é evidenciada pelo fenótipo de humanos com deficiências no regulador autoimune (AIRE, do inglês autoimmune regulator) molecular. O AIRE é expresso por células estromais medulares tímicas e estimula a expressão de autoantígenos, em especial aqueles que são expressos normalmente nas glândulas endócrinas. A doença resultante das mutações no gene do AIRE, poliendocrinopatia autoimune-candidíase-distrofia ectodérmica (APECED, do inglês autoimmune polyendocrinopathy-candidiasis-ectodermal dystrophy), é causada pela grande abundância de células T autorreativas que não passaram por uma seleção negativa apropriada.
Apesar da presença de mecanismos normais de tolerância central, algumas células autorreativas podem escapar da seleção negativa que ocorre na medula óssea e no timo e penetrar nos órgãos linfoides periféricos e na circulação. Algumas células T autorreativas que escapam da seleção negativa no timo são recrutadas em um pool de células T reguladoras naturais (nTreg) com funções supressivas imunes. Essas células nTreg, um subgrupo as células Th, poderão ser distinguidas pela expressão do fator de transcrição gênica FoxP3, que é essencial para as funções reguladoras. As mutações de sentido incorreto no lócus do gene FOXP3 resultam nas síndromes humanas IPEX (imune dysregulation, polyendocrinopathy, enteropathy, X-linked disease [desregulação imune, poliendocrinopatia, enteropatia, doença ligada ao X]) e XLAAD (X-linked autoimmunity-allergic dysregulation syndrome [autoimunidade ligada ao X-síndrome da desregulação alérgica]) que, com frequência, são fatais em idades precoces. Nos anos mais recentes, houve também um interesse crescente em populações de células B com propriedades reguladoras imunes. Os processos que resultam do desenvolvimento de células B reguladoras (Breg) e, em especial, se elas são preferencialmente autorreativas, ainda não são muito bem compreendidos.

Figura 1 - O reconhecimento de antígenos (Ag)-complexo de histocompatibilidade principal (MHC) pelo receptor de células controla a sobrevivência e a ativação das células T. Os Ags são processados no interior das células apresentadoras de antígenos e são apresentados na superfície celular por moléculas do MHC. O lócus do gene do MHC é uma herança do pai e da mãe e, além disso, é um sítio altamente polimórfico. Isso estabelece um amplo repertório de diversidades genéticas dentro da população e diminui a probabilidade de que dois indivíduos, a não ser gêmeos idênticos, apresentem exatamente o mesmo fenótipo de MHC. A estrutura do sulco de ligação da molécula de MHC restringe a forma e as propriedades moleculares dos antígenos que poderão ser apresentados por aquele MHC. O receptor de células T (TCR) liga-se ao MHC e ao complexo Ag com afinidades diferentes, de acordo com número de pontos de contato e de forças moleculares, como as interações hidrofóbicas e iônicas. A força da interação entre o TCR e o Ag-MHC pode exercer influências diferentes sobre o resultado, dependendo da localização anatômica e do estado evolucionário da célula T. A diversidade do TCR tem origem em recombinações genéticas, porém cada célula T expressa apenas um único TCR recombinado. (a) As recombinações de TCR não produtivo resultam na falha de seleção positiva no timo e na deleção daquela célula T antes de sua penetração na periferia. (b) As interações de baixa afinidade durante a seleção tímica favorecem a sobrevivência da célula T, permitindo sua passagem para a periferia. O reconhecimento antigênico fraco pelo TCR na periferia resulta em nenhuma ativação ou em uma ativação baixa. (c) As interações de alta afinidade do TCR no timo levam à morte celular induzida por ativação ou ao desvio no subgrupo regulador, enquanto estiver na periferia; o reconhecimento antigênico forte leva à ativação e à expansão clonal da célula T respondente. A figura não mostra a interação entre as moléculas CD8 e CD4 da célula T e a porção externa das moléculas de MHC de classe I e classe II, respectivamente. Essa interação acessória no timo resulta no comprometimento da linhagem da célula T em células T auxiliares CD4+ ou em linfócitos T citotóxicos CD8+ com funções efetoras distintas.
Existem mecanismos adicionais de seleção negativa que agem sobre linfócitos mais maduros e que mantêm a tolerância imune periférica. Esses processos ocorrem em órgãos linfoides secundários, em sítios de privilégio imune (olhos e órgãos reprodutivos) e em tecidos inflamados. A mediação da tolerância periférica é feita principalmente através da morte celular programada (apoptose) por meio da interação entre os receptores específicos de morte localizados na célula alvo e seus ligantes indutores de morte, expressos por células destruidoras ou citotóxicas especiais. As mutações de perda funcional em um desses receptores de morte – Fas – ou em seu parceiro de ligação – ligante Fas – aumenta a sobrevida dos linfócitos periféricos e da síndrome linfoproliferativa autoimune (SLPA). A apoptose de linfócitos periféricos depende da ativação dos respectivos receptores de antígenos e, consequentemente, é um mecanismo que controla não apenas os linfócitos T e B autorreativos periféricos, mas também os linfócitos cujos alvos são microrganismos infecciosos e outros antígenos.
Nas reações imunes típicas a algum microrganismo patogênico recentemente encontrado, a primeira resposta é mediada pelo sistema imune inato. Essa mediação poderá ser induzida por interações diretas entre as células imunes inatas e seus receptores de reconhecimento de padrões (PRRs, do inglês pattern recognition receptors) e os micróbios no tecido afetado, ou poderá ser iniciada indiretamente através de danos teciduais causados pelos efeitos do micróbio sobre as células não imunes que, em seguida, ativam a imunidade inata por meio da liberação de “sinais de alerta”. Em qualquer um dos casos, a resposta inflamatória disparada liberará os fatores solúveis envolvidos nas comunicações intercelulares, em particular as citocinas pró-inflamatórias e as citocinas quimioatrativas (quimiocinas). Os papeis imediatos dessas citocinas pró-inflamatórias e das quimiocinas são ações locais e sistêmicas para induzir mecanismos de defesa de hospedeiro que lentificam e neutralizam a patogenicidade dos micróbios. Muitos dos efeitos desses mediadores se manifestam em sinais clínicos que são indicativos de infecção e inflamação, febre, edema, eritema, choque e elevações proteicas na fase aguda.
Um exemplo de resposta pró-inflamatória é a situação em que os macrófagos teciduais detectam a presença de uma infecção bacteriana através da ativação do receptor toll-like (TLR) e liberam citocinas, tais como o fator a de necrose tumoral (TNF-a, do inglês tumor necrosis factor alpha); a interleucina 1ß (IL-1ß), IL-6 e IL-12; e a quimiocina CXCL8. A TNF-a, IL-1ß e IL-6 têm muitos efeitos colaborativos, incluindo indução local de permeabilidade vascular e expressão de moléculas de adesão celular, que facilitam o fluxo interno de outras células e mediadores imunes no sítio infectado. Essas citocinas induzem também a mobilização de neutrófilos da medula óssea através de uma ação sistêmica para elevar a temperatura do corpo por meio da estimulação do hipotálamo e pelo aumento na produção de energia nos tecidos adiposos e musculares, assim como para ativar a resposta de fase aguda no fígado. A resposta de fase aguda, que é particularmente sensível a níveis elevados de IL-6, aumenta a produção de pentraxinas (proteína C reativa e proteína amiloide sérica), de lectina de ligação a manose e de outras moléculas envolvidas na ativação de complementos. A IL-6 age também sobre os linfócitos para aumentar a produção de anticorpos e influenciar a ativação e diferenciação de células Th. Um dos resultados da liberação de anticorpos induzida por inflamação, de reagentes da fase aguda e de complementos, é revestir a superfície das bactérias, processo conhecido por opsonização, que facilita o domínio do patógeno por macrófagos, células dendríticas e de neutrófilos por fagocitose. A IL-12 estimula a ativação de células destruidoras naturais e influencia a diferenciação de células Th. O CXCLS age como se fora um sinal de busca para o recrutamento de leucócitos polimorfonucleares (PMNs, do inglês polymorphonuclear leukocytes) no interior do sítio da infecção. Consequentemente, os diversos efeitos desse conjunto de mediadores pró-inflamatórios lentificam o crescimento das bactérias, de forma cooperativa, facilitando a fagocitose e iniciando a ativação do sistema imune adaptativo, de acordo com a necessidade.
As infecções bacterianas localizadas e a resposta imune inata resultante de macrófagos teciduais são apenas um entre muitos mecanismos possíveis para indução da imunidade inata. Outros patógenos (vírus, fungos, vermes parasitários) exigem tipos distintos de respostas imunes para seu controle e, portanto, desencadeiam tipos diferentes de inflamação por meio do reconhecimento de seus antígenos específicos. Além disso, a extensão da infecção e sua localização anatômica são fatores importantes que afetam o tipo de células envolvidas na resposta inicial e os mediadores que são produzidos.
A resposta inicial às infecções virais compartilham algumas características sobrepostas com as infecções bacterianas que envolvem outros tipos de células e de mediadores, que são mais eficientes para induzir a resposta imune citotóxica necessária para eliminar as células hospedeiras que abrigam vírus. É importante observar que as células efetoras inatas principais formam um subgrupo de células dendríticas plasmacitoides (pDCs), sendo que a estimulação dos respectivos PRRs predominantes por antígenos virais resulta principalmente na produção de grandes quantidades de citocinas interferon tipo 1, interferon alfa e interferon beta. Os interferons inibem a replicação viral; induzem a expressão de moléculas do MHC; estimulam a produção das quimiocinas CXCL9, CXCL10 e CXCL11 (quimiocinas especializadas na atração de células que têm como alvo as infecções virais); e ativam linfócitos T citotóxicos (LTCs) CD8+ que são especializados na destruição de células infectadas por vírus. Por outro lado, os vermes helmintos parasitas e os respectivos ovos são organismos multicelulares de grande porte resistentes à fagocitose e à ação de células citotóxicas. Esses organismos estimulam a produção de IL-4 a partir de mastócitos, basófilos e linfócitos, que orientam o início de respostas imunes adaptativas que favorecem a expulsão dos vermes.
Em alguns casos, a resposta imune inata é suficiente para eliminar a ameaça, sendo desnecessário disparar a imunidade adaptativa. Portanto, provavelmente a frequência de infecções de baixo nível e de respostas inflamatórias subclínicas seja bem mais elevada do que se imagina. A diferença entre aquilo que é considerado um microrganismo patogênico e um microrganismo benigno possivelmente não reflita sua capacidade para infeccionar ou inibir os indivíduos, porém, em vez disso, talvez seja mais dependente da extensão pela qual é capaz de ativar e/ou subverter defesas imunes existentes no hospedeiro, resultando em doença clínica. Entretanto, essas infecções de baixo nível podem influenciar a homeostase no sistema imune e afetar as respostas na direção de encontros subsequentes com microrganismos patogênicos ou a tolerância imune a autoantígenos.
É necessário estimular respostas imunes adaptativas nas situações em que as respostas imunes inatas iniciais a uma infecção forem insuficientes para controlar os patógenos. Conforme mencionamos acima, as respostas imunes adaptativas são direcionadas por células T auxiliares que têm capacidade para integrar e interpretar os sinais provenientes do microambiente. Em seguida, as células T auxiliares produzem conjuntos específicos de superfícies celulares e de mediadores solúveis que enviam sinais para outras células para que possam montar respostas apropriadas e intensificadas para estimular os patógenos. Os efeitos devastadores do HIV, que infecta as células T CD4+, ilustram a importância dessas células para abrigar defesas.
O primeiro sinal para ativação de uma célula T auxiliar periférica envolve o reconhecimento de seu antígeno específico apresentado no contexto de uma molécula de MHC de classe II na superfície de uma célula apresentadora de antígeno (APC, do inglês antigen-presenting cell). Entre essas populações de células imunes inatas, as células dendríticas, os macrófagos ativados e os linfócitos B-1 possuem capacidades especiais de dominar os microrganismos e de processar suas proteínas em fragmentos de peptídeos que se ligam ao MHC de classe II e são transportados para a superfície celular. Os linfócitos B-2 que forem ativados por meio da ligação do antígeno ao receptor de células B também são capazes de receber antígenos e processá-los em MHCs de classe II. Essas populações de APC “profissional” interagem com as células T auxiliares, principalmente nos órgãos linfoides secundários, e geram não apenas o primeiro sinal para ativação das células T auxiliares através da apresentação de antígenos, mas também sinais acessórios, que se denominam coestimulação, na forma de ligantes expressos nas superfícies celulares e nas citocinas solúveis. Essas moléculas coestimuladoras de sinalização são reguladas por respostas imunes inatas e incluem algumas das citocinas que foram mencionadas acima.
Acredita-se que a coestimulação por APCs gera sinais através dos quais as células T auxiliares conseguem sobreviver e proliferar. Além disso, a natureza dos agentes estimuladores se reflete no tipo específico de coestimulação fornecido pela célula apresentadora de antígeno que, por sua vez, controla o conjunto correto de moléculas e de células efetoras que foram mobilizadas durante a resposta adaptativa da célula T. Na ausência de coestimulação, a ativação de células T auxiliares virgens através do receptor de células T resulta na desativação funcional (anergia) ou na morte da célula T auxiliar. Um exemplo importante de coestimulação que ocorre na maior parte dos encontros de células T virgens produtivas/células apresentadoras de antígenos é a interação dos membros B7.1 (CD80) e B7.2 (CD86) da família B7 com o receptor de CD28 expresso nas células T auxiliares virgens. O segundo sinal gerado por essa interação, em combinação com o reconhecimento de antígenos por receptores de células T, induz a produção de IL-2, o principal fator de crescimento de células T auxiliares, assim como o aumento na expressão do receptor de IL-2. Consequentemente, a liberação de IL-2 por células T auxiliares ativadas age de uma forma autócrina para estimular a proliferação de células T. Outro exemplo de coestimulação geralmente ocorre entre as células T e as células apresentadoras de antígenos, seja qual for o antígeno resultante, é a interação do antígeno 1 (LFA-1) (CD11a/CD18) associado à função de linfócitos da molécula de integrina da célula T dimérica com os respectivos ligantes, como a molécula-1 de adesão intercelular (ICAM-1, do inglês intercellular adhesion molecule-1) (CD54). Essas moléculas podem iniciar a interação antes do envolvimento dos receptores de células T (TCR, do inglês T cell receptor); no entanto, as alterações conformacionais para LFA-1, após envolvimento do TCR, prolongam a interação entre as duas células e intensificam a ativação das células T auxiliares. As expressões de B7 e de ICAM nas APCs são suprarreguladas por citocinas pró-inflamatórias, demonstrando a presença de outro mecanismo de comunicação cruzada entre a imunidade inata e a imunidade adaptativa. Esses são apenas dois exemplos de um conjunto complexo de interações coestimuladoras que ocorrem entre células T auxiliares e células apresentadoras de antígenos. Muitas das outras interações apresentam efeitos de sobreposição e efeitos redundantes, comparáveis àqueles que foram descritos acima, e geralmente servem para ativar as células T auxiliares sem desviar suas respostas. Outras interações de moléculas coestimuladoras direcionam as células T auxiliares para diferenciar caminhos efetores mais específicos.
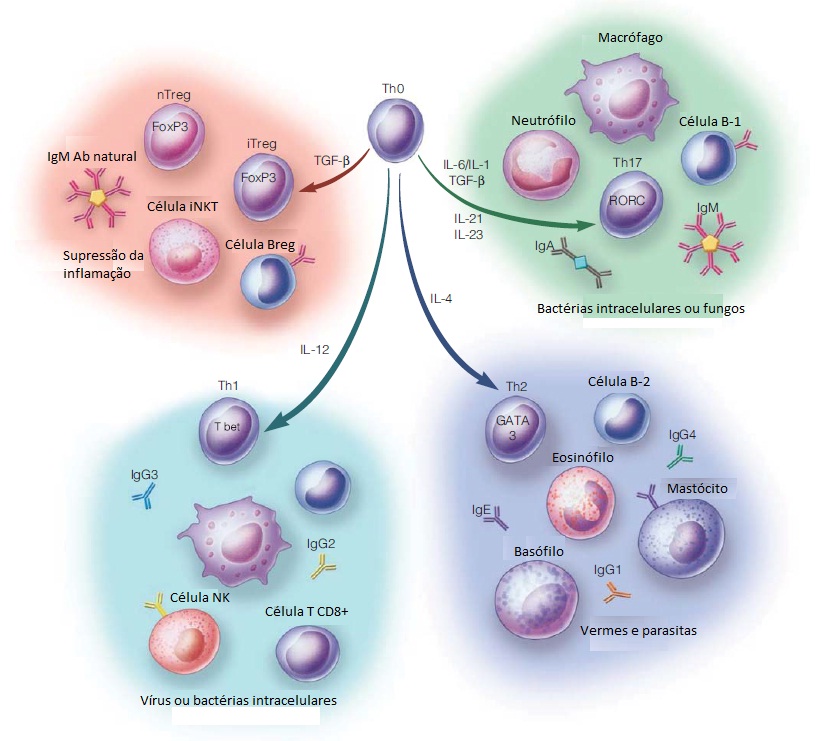
Patógenos intracelulares como vírus e alguns tipos de bactérias exigem respostas de hipersensibilidade do tipo mediada ou retardada para transmitir instruções para as células infectadas para eliminar o patógeno ou sofrer apoptose. Essa resposta é direcionada por células T auxiliares que foram diferenciadas em células produtoras de interferon gama [ver a Figura 2]. Essas células são designadas células Th1 e são distinguidas mais conclusivamente de outros subgrupos de células Th pela expressão do fator de transcrição T-bet, que faz a mediação da expressão dos genes específicos da célula Th1. Elas têm origem nas células T auxiliares (células Th) virgens que são coestimuladas pela IL-12 no momento da apresentação do antígeno. O interferon gama tem muitos efeitos sobre as células do sistema imune, incluindo estimulação da atividade microbicida celular intrínseca em macrófagos, indução da expressão de MHC de classe I e classe II, intensificação das vias processadoras de antígenos, ativação de linfócitos T citotóxicos e de células destruidoras naturais, e estimulação da mudança de isotipos em linfócitos B-2 para a produção das subclasses IgG2 e IgG3. Além disso, o interferon gama promove a diferenciação ulterior das células Th virgens na linhagem Th1 e exerce efeitos supressivos nos subconjuntos de células Th que produzem outros padrões de citocinas.
As respostas imunes inatas contra patógenos extracelulares favorecem a indução de imunidade humoral que, em um sentido amplo, se refere a reações imunes dominadas por anticorpos e complementos derivados de células B-2. Dentro dessa classificação estão os subgrupos de células Th que se especializam na defesa contra parasitas, fungos e bactérias extracelulares. A presença de IL-4 fúngica ou induzida por vermes neste momento da apresentação antigênica é o sinal principal para diferenciação de células Th virgens na linhagem de células Th2 [ver a Figura 2]. O fator de transcrição GATA-3 é imprescindível para a diferenciação de células Th2. As células Th2 produzem principalmente as citocinas IL-4, IL-13 e IL-5. Assim como o interferon gama no interior da linhagem de células Th1. A IL-4 promove a expansão das células Th2 e suprime outras linhagens de células Th. A IL-4 é também um fator de crescimento importante para os linfócitos B-2 e promove a troca de isotipos de anticorpos nas células B-2 para a produção das subclasses de anticorpos IgG1, IgG4 e IgE. Os anticorpos IgE são particularmente importantes na reação contra vermes parasitas porque se ligam a receptores Fce específicos nas superfícies de mastócitos, basófilos e eosinófilos e servem de mediadores no processo de degranulação a partir dessas células. A IL-13 tem muitos efeitos sobrepostos com a IL-4 e está envolvida na supressão da ativação de macrófagos e na indução de fibrose. A IL-5 é um fator de crescimento e de diferenciação de eosinófilos.
Em 2005, a linhagem Th17 das células Th foi diferenciada fenotípica e funcionalmente das células Th1 e, a partir de então, vem sendo investigada intensamente por causa de seu papel patogênico nas doenças autoimunes. Uma das características diferenciadoras das células Th17 é a expressão do fator de transcrição RORC. A diferenciação das células Th17 é acionada pela coestimulação com citocinas pró-inflamatórias (principalmente IL-6 ou IL-1ß) em combinação com o fator de crescimento transformador ß (TGF-ß, do inglês, transforming growth factor ß). Ulteriormente, a diferenciação das células Th17 é promovida pelos efeitos sequenciais de IL-21 e IL-23 e fortemente inibida pela presença de interferon gama ou de IL-4. Aparentemente, as funções normais das células Th17, que são mediadas pelas citocinas características IL-17A, IL-17F e IL-22, têm como foco principal promover informações e dar proteção contra a ação de patógenos extracelulares nas superfícies mucosas, incluindo leveduras e bactérias. As interleucinas IL-17A e IL-17F induzem a expressão de citocinas e quimiocinas por células não linfoides, resultando no recrutamento de neutrófilos para os sítios de inflamação. A interleucina IL-22 promove as funções de barreira das células epiteliais do revestimento de mucosas, além de ser outro indutor de reações de fase aguda que promove a ativação de complementos.
Outra via importante de diferenciação de células Th ativadas leva aos subconjuntos de células T reguladoras (Treg). As células nTreg foram introduzidas acima como um resultado para as células Th autorreativas que escaparam da seleção tímica negativa. As células induzidas ou iTreg resultam da ativação de células T periféricas na presença de co-estimulação pelo TGF-ß e na ausência de citocinas pró-inflamatórias. Tanto as células nTreg como as células iTreg são marcadas pela expressão do fator de transcrição FoxP3 e compartilham a produção das citocinas efetoras principais TGF-ß e IL-10. Além do papel de promover a diferenciação das células Th17 e iTreg, o TGF-ß possui efeitos anti-inflamatórios potentes através do bloqueio da divisão de células T e da produção de citocinas, assim como através da indução de apoptose em células imunes. O TGF-ß possui também efeitos estimuladores sobre a indução de isotipos de anticorpos mudando para IgA, que é segregado nas superfícies mucosas, além de ser muito importante para neutralizar patógenos associados a mucosas. A IL-10 também tem efeitos inibidores sobre a produção de citocinas de células T, em particular IL-2, TNF-a e IL-5. A IL-10 suprime a ativação de macrófagos e de células dendríticas e a expressão de IL-2 e MHC e de moléculas coestimuladoras por células apresentadoras de antígenos (APCs, do inglês antigen-presenting cells), limitando, consequentemente, suas capacidades de apresentação de antígenos. Os efeitos positivos da IL-10 sobre o sistema imune incluem promoção da sobrevivência e maturação de células B em células plasmáticas produtoras de anticorpos altamente ativas.
Outro mecanismo importante da supressão imune por células Treg é a expressão superficial do antígeno-4 associado ao linfócito T citotóxico molecular (CTLA-4). Sob o ponto de vista estrutural, o CTLA-4 está relacionado ao receptor CD28, que é um coestimulador de células T e se liga a B71 e B7.2, embora com uma afinidade muito maior do que CD28. Ao contrário do CD28, a ligação de CTLA-4 envia um sinal inibidor para as células Th e bloqueia a ativação mediada por TCR. A expressão superficial de CTLA-4 é suprarregulada nas células Th após a ativação e desempenha um papel importante na regulação imune. Esse fato é evidenciado pelo fenótipo de camundongos com deficiência grave de CTLA-4, que apresentam níveis elevados de proliferação de células T em múltiplos órgãos e de mortalidade na juventude. O papel do CTLA-4 na função das células Treg pode ser mediado indiretamente por meio da interação das células Treg com as células apresentadoras de antígenos.
Consequentemente, o sistema imune saudável é um conjunto bem afinado e altamente complexo de células e de mediadores solúveis que agem rapidamente para controlar muitas ameaças comuns, embora seja também altamente flexível e adaptável para combater os ambientes microbianos em constante mutação onde vivem os seres humanos. Uma das consequências da flexibilidade é que surgem especificidades indesejadas no repertório imune, o que exige um sistema potente de verificações e de balanceamentos para manter a homeostase e a tolerância aos autoantígenos. Nas seções seguintes mudaremos o foco para os erros cometidos pelo sistema imune que contribuem para as doenças humanas, assim como os tratamentos atuais e prospectivos de distúrbios imunomediados.
Figura 2 - As células T auxiliares (Th) são determinantes centrais das respostas imunes adaptativas em relação a estímulos diferentes. Os linfócitos T auxiliares (Th0) virgens são capazes de processar os sinais provenientes da superfície de células apresentadoras de antígenos, assim como as citocinas mostradas na figura. A integração desses sinais resulta no comprometimento das células Th com um fenótipo efetor que é expresso pela expressão de fatores distintos de transcrição gênica (apresentados nos núcleos das células Th diferenciadas). Cada célula efetora do tipo Th produz citocinas que promovem a ativação de outros tipos de células e a produção de isotipos dos anticorpos apresentados na figura. Alguns dos anticorpos produzidos se ligam aos receptores Fc nas células do sistema imune inato, agindo como mecanismo receptor adicional para gerar as funções efetoras das células imunes inatas. AC = anticorpo; Breg (regulatory B cell) = célula B reguladora; IL = interleucina; iNKT (invariant natural killer T cell) = célula T destruidora natural invariante; iTreg (induced regulatory T cell) = célula T reguladora induzida; NK (natural killer) = destruidora natural; nTreg (natural regulatory T cell) = célula T reguladora natural; TGF-ß (transforming growth factor-ß) = fator-ß transformador do crescimento; Th (T helper) = auxiliar T; Treg (regulatory T cell) = célula T reguladora.
O tratamento das doenças autoimunes deve se fundamentar na avaliação dos mecanismos patogênicos essenciais para as manifestações clínicas de uma doença específica. Por exemplo, a patogênese de artrite reumatoide (AR) é uma interação complexa entre o sistema imune inato e o sistema imune adaptativo; portanto, AR é uma condição em que múltiplas formas de modulação ou supressão imune são eficazes. A sobreposição desses mecanismos patogênicos é apenas parcial com os mecanismos presentes em outras doenças autoimunes comuns, como o lúpus eritematoso sistêmico (LES). Consequentemente, algumas abordagens utilizadas no tratamento de AR também são eficazes no tratamento de LES, enquanto que outras abordagens poderão agravar o LES. O uso crescente de agentes biológicos específicos no tratamento de autoimunidades está ampliando os conhecimentos sobre a patogênese da doença através do sucesso terapêutico e de toxicidades inesperadas. A complexidade das respostas imunes se reflete na diversidade das doenças autoimunes e de seus mecanismos patogênicos. Algumas condições inflamatórias são totalmente acionadas por mecanismos que em geral estão envolvidos em respostas imunes inatas, sem evidências de alterações na imunidade adaptativa. Essas condições, incluindo as síndromes de febre periódica, são chamadas apropriadamente de “doenças autoinflamatórias”.
Tipicamente as doenças autoimunes se caracterizam pela formação de anticorpos patogênicos e/ou pela ativação de linfócitos T autorreativos. Conforme discutimos acima, as respostas imunes humorais e as celulares sofrem uma regulação cruzada; portanto, as doenças mais autoimunes envolvem a ativação de células Th e de células B, ainda que os mecanismos patogênicos sejam altamente focados em um tipo ou outro de resposta. Os autoanticorpos podem provocar doenças clínicas através de uma vasta gama de mecanismos, alguns dos quais são ilustrados na Tabela 2. Em alguns exemplos de doenças mediadas por autoanticorpos, como doença tireoidea autoimune e miastenia grave, os autoantígenos são bem definidos e são específicos de órgãos. A patogênese é mediada diretamente pela ligação de anticorpos, resultando na ativação inapropriada do receptor alvo, como na doença de Graves, ou no bloqueio da ligação do receptor com seu ligante funcional ou na desativação, como nos casos de miastenia grave. Em outras condições com domínio de anticorpos, como lúpus eritematoso sistêmico (LES), a faixa de autoantígenos é mais diversificada e envolve uma ampla gama de tecidos e tipos de células. A patogênese de LES envolve a ativação de complementos e a formação de complexos imunes que podem ser depositados em tecidos e ativar respostas inflamatórias. A ligação de autoanticorpos poderá causar também a destruição de células ou tecidos, como nos casos de anemia hemolítica autoimune. Apesar da definição de mecanismos patogênicos em algumas doenças, ainda falta delinear o papel exato dos autoanticorpos em várias outras doenças autoimunes. Os exemplos incluem anticorpos para proteínas contendo citrulina, que estão intimamente ligadas à artrite reumatoide, e anticorpos para enzimas neutrofílicas em vasculites associadas ao anticorpo anticitoplásmico de neutrófilos (ANCA, do inglês antineutrophil cytoplasmic antibody). Nem todos os autoanticorpos são necessariamente patogênicos. Alguns autoanticorpos podem surgir como consequência ao invés de causa. Com frequência, é comum a presença de autoanticorpos de baixa titulação em indivíduos saudáveis, sendo que há evidências crescentes que dão suporte ao conceito de que alguns tipos de anticorpos autorreativos, incluindo anticorpos IgM “naturais” em circulação e anticorpos IgG sialilados, desempenham papeis imunes supressivos e protetores nas doenças autoimunes.5,6
Tipicamente, doenças autoimunes mediadas por células, tais como artrite reumatoide (AR), esclerose múltipla (EM) e diabetes tipo 1 possuem um componente genético que incluem ligação a um subgrupo de alelos da molécula de classe II do complexo de histocompatibilidade principal (MHC, do inglês major histocompatibility complex) altamente polimórfico. Esse fato sugere que os autoantígenos específicos de órgãos apresentados por esses alelos do MHC acionam respostas imunes mediadas por células Th. Entretanto, o fato de que muitos indivíduos na população que têm alelos do MHC associados a alguma doença e, mesmo assim, nunca apresentam sintomas, indica que são necessários conhecimentos adicionais para esclarecer o(s) mecanismo(s) exato(s) através dos quais os alelos específicos do MHC se associam a várias doenças autoimunes. O envolvimento comprovado de linfócitos T e B na patogênese de doenças autoimunes resultou no desenvolvimento de novas estratégias terapêuticas com foco na regulação de suas funções.
As células B produzem autoanticorpos, agem como células apresentadoras de antígenos (APCs, do inglês antigen-presenting cells), segregam citocinas pró-inflamatórias como a IL-16 e podem regular as células T. Portanto, a depleção de células B possui potencial terapêutico em muitos níveis da patogênese autoimune. O rituximab, um anticorpo monoclonal quimérico direcionado contra o antígeno CD20, é expresso na superfície de linfócitos B, excetuando-se os precursores precoces de células B e células plasmáticas diferenciadas. O rituximab é uma imunoglobulina IgG1 kappa que recruta células efetoras para promover a lise de linfócitos B por meio da citotoxicidade mediada por células dependente de complemento ou citotoxicidade celular dependente de anticorpos (CCDA). A citotoxicidade mediada por células dependente de complemento envolve a interação entre o sistema imune inato e o sistema imune adaptativo. O componente do complemento – C1q – interage com regiões Fc de anticorpos ligados a antígenos alvos levando à ativação das enzimas C1r e C1s, seguida pela clivagem de C4 e C2 e desencadeando a rota clássica de ativação de complementos. Ao longo desse processo, há o reconhecimento de C3b e C4b pelas células imunes efetoras através dos respectivos receptores de complementos, intensificando a fagocitose e a citotoxicidade.
|
Tabela 2: Doenças Humanas Mediadas por Autoanticorpos: Exemplos de Mecanismos Patogênicos
| ||
|
Doença |
Alvo Molecular/Celular |
Mecanismo Patogênico
|
|
Doença de Graves |
Receptor do hormônio estimulante da tireoide (TSH)
|
Agonista de receptores |
|
Miastenia grave |
Receptor da acetilcolina |
Bloqueio de receptores
|
|
Nefrite causada por LES |
DNA, antígenos nucleares |
Deposição do complexo imune
|
|
Anemia hemolítica autoimune |
Eritrócitos |
Fagocitose e citotoxicidade celular mediada por complementos
|
|
Síndrome antifosfoslipídica |
Glicoproteina ß2 1 |
Promoção de trombose
|
LES = lúpus eritematoso sistêmico; TSH (thyroid stimulating hormone) = hormônio estimulante da tireoide.
A CCDA é mediada por células NK que expressam CD16, um receptor de FcyRIII que reconhece a porção Fc dos anticorpos IgG1 ou IgG3 que se ligam à superfície de células-alvo. Assim que o receptor Fc de uma célula NK é acoplado pela região Fc do IgG, a célula Nk libera citocinas como interferon gama e grânulos citotóxicos contendo perforina e granzimas no interior da célula alvo, promovendo a morte celular ao desencadear a apoptose. Esses mecanismos de lise celular são pertinentes ao mecanismo de algum dos outros anticorpos monoclonais que são utilizados no tratamento de doenças imunes.7 Considerando que a molécula CD20 não é expressa por células-tronco da medula óssea, por precursores precoces de células B ou por células plasmáticas diferenciadas, a repopulação do compartimento de células B ocorre gradualmente, logo após a eliminação do rituximab.8 O rituximab é eficaz no tratamento de AR e foi aprovado para essa aplicação, embora não tenha sido aprovado para tratamento de lúpus eritematoso sistêmico (LES).
Provavelmente, as células Th17 estejam envolvidas logo no início da patogênese de uma grande variedade de doenças autoimunes e inflamatórias, incluindo AR, doença intestinal inflamatória, esclerose múltipla e outras condições.9 Há um número crescente de células Th17 e níveis anormalmente elevados de citocinas IL-6 e IL-23 indutivas de Th17 detectadas no sangue periférico de pacientes com artrite reumatoide, em comparação com indivíduos dos grupos de controle.10,11 Nos casos de AR, a IL-17 produzida por células Th17 ativa fibroblastos sinoviais, monócitos, macrófagos e condrócitos levando à produção de IL-6, IL-8 e de enzimas destruidoras de tecidos como as metaloproteinases matriciais.12
A diferenciação das células Th17 nos seres humanos não é completamente caracterizada, embora a IL-6 promova a diferenciação de Th17 in vitro, sendo que a IL-23 e a IL-1ß também desempenham papeis importantes nesse processo. Além disso, a IL-6 suprime o desenvolvimento de células T virgens em células Treg. Não há nenhum medicamento aprovado pela Food and Drug Administration (FDA) com alvo direto em IL-17 ou nas células Th17; no entanto, o bloqueio de IL-6 poderia resultar em inflamações induzidas pela redução de IL-6 e em alterações no equilíbrio da diferenciação Th17/Treg, suprimindo o desenvolvimento e as funções das células Th17.13 O tocilizumab é um anticorpo IgG1 monoclonal kappa humanizado que age contra o receptor da IL-6 que pode bloquear a ação da interleucina-6 e foi aprovado pela FDA para uso no tratamento de artrite reumatoide.
A IL-23, uma das citocinas presentes em lesões cutâneas causadas pela psoríase, dá suporte ao desenvolvimento, expansão e ativação das células Th17.14 O ustekinumab é um anticorpo monoclonal direcionado contra a subunidade p40 compartilhada por IL-12/IL-23 com alta afinidade e especificidade que, consequentemente, evita a interação entre essas duas citocinas e seu receptor celular superficial. Ao diminuir a atividade biológica da IL-23 e da IL-12, o ustekinumab poderia afetar negativamente a diferenciação de Th17 e Th1 e a diferenciação da IL-17 secretora de células CD8+ presentes nas placas cutâneas psoriáticas. O ustekinumab é altamente eficaz nos casos de psoríase e foi aprovado pela FDA para essa indicação.
Outros loci de genes, incluindo vários genes que codificam as proteínas envolvidas no desenvolvimento e na função de linfócitos T e B, estão associados a doenças autoimunes como artrite reumatoide, lúpus eritematoso sistêmico e esclerose múltipla. A identificação desses genes esclareceu novos mecanismos patogênicos que poderão estar envolvidos no início e/ou na manutenção da autoimunidade mediada por células Th e da autoimunidade mediada por anticorpos, assim como a identificação de novos alvos terapêuticos. Além dos alvos imunes adaptativos, os novos conhecimentos sobre as funções aberrantes do sistema imune inato na autoimunidade, em especial no nível de apresentação de antígenos, estão lançando alguma luz nos circuitos reguladores cruzados envolvidos no controle da imunidade adaptativa e na prevenção de doenças autoimunes.
O sistema imune inato desempenha um papel importante na artrite reumatoide (AR) evidenciado pela presença de níveis elevados de citocinas derivadas de macrófagos, TNF-a, IL-1 e IL-6 no soro e no sinóvio.15 Os macrófagos servem de biomarcadores para doenças ativas, levando-se em consideração que qualquer redução no número de macrófagos no sub-revestimento dos tecidos sinoviais se correlaciona com o grau de melhora clínica obtida com uma multiplicidade de terapias escolhidas para o tratamento de artrite reumatoide.16 Os medicamentos usados com mais frequência no tratamento de AR – prednisona, metotrexato, leflunomida, sulfassalazina e inibidores do TNF-a – reduzem o número de macrófagos no revestimento sinovial e melhoram os resultados clínicos da doença.17 Os inibidores de citocinas, incluindo os inibidores do TNF-a (adalimumab, certolizumab pegol, etanercept, golimumab, infliximab), os antagonistas de receptores de interleucina-1 (IL-1RA) (anakinra) e o anticorpo do receptor da IL-6 (tocilizumab) têm como alvo as citocinas produzidas por macrófagos sinoviais.
O TNF-a solúvel e seu precursor TNF-a transmembranar têm algum envolvimento nas respostas inflamatórias. A expressão do precursor TNF-a transmembranar ocorre em macrófagos e linfócitos ativados. A clivagem da forma solúvel de TNF-a ocorre a partir da forma transmembranar por meio da enzima conversora do fator de necrose tumoral alfa (TACE, do inglês tumor necrosis factor-a converting enzyme). O TNF-a solúvel se liga aos receptores 1 e 2 do TNF para mediar os efeitos biológicos sobre os tecidos mais distantes.7 O TNF-a transmembranar se liga aos receptores 1 e 2 do TNF nas células-alvo, liberando um sinal para as células-alvo e um sinal inverso no interior do TNF-a transmembranar contendo células.18
Os agentes anti-TNF infliximab, adalimumab e etanercept ligam e neutralizam o TNF-a solúvel, porém produzem efeitos distintos sobre a forma transmembranar do TNF-a. O infliximab, a adalimumab e o etanercept contêm os domínios Fc da IgG1 envolvida na ligação dos receptores Fc nas células NK, facilitando a lise de células-alvo por citotoxicidade celular dependente de anticorpos (CCDA).7 O infliximab e o adalimumab demonstraram atividade CCDA mais efetiva usando células-alvo transformadas, em comparação com o etanercept, embora todos os três sejam semelhantes em relação à atividade CCDA direcionada para as células T transfectadas do TNF-a transmembranar.19-21 O certolizumab pegol não apresentou nenhuma atividade CCDA, provavelmente devido à ausência de domínios Fc de IgG1.21 Presumivelmente, o golimumab tenha atividade CCDA, tendo em vista que possui domínios Fc de IgG1.
Os receptores de reconhecimento de padrões (PRRs, do inglês pattern recognition receptors) denominados NALPs (domínios NACHT [neuronal apoptosis inhibitor protein], proteína inibidora da apoptose neuronal; repetição rica em leucina [LRR, do inglês leucine-rich repeat] e domínio da pirina [PYD, do inglês pyrin domain]) se localizam nos citoplasmas de células imunes inatas como os sinoviócitos semelhantes a fibroblastos, que residem no revestimento articular. Os NALPs formam uma estrutura denominada inflamassomo que responde aos sinais perigosos incluindo componentes patogênicos ou, nos casos de gota, cristais de urato monossódico. A criopirina é um dos membros da família de NALPs que se expressa no sinóvio e é induzível pelo TNF-a.22 Em resposta aos sinais de alerta, o inflamassomo processa IL-1 em sua forma ativa através do envolvimento da caspase-1.23
Os níveis de IL-1RA que ocorrem naturalmente no sinóvio de artrite reumatoide e no líquido sinovial não são suficientes para competir com a IL-1 produzida na articulação. O medicamento anakinra é uma forma de IL-1RA humana recombinante não glicosilada que inibe competitivamente a ligação de IL-1 ao receptor tipo 1 da interleucina-1. Consequentemente, o uso de anakinra possivelmente bloqueie os efeitos do excesso de IL-1. Embora tenha sido aprovado pela FDA para tratamento de artrite reumatoide, o medicamento anakinra raramente é usado no gerenciamento da doença, levando-se em conta que os outros agentes são mais eficazes. Esse tipo de medicamento é mais útil no tratamento de condições autoinflamatórias.
As células dendríticas, assim como os macrófagos e as células B, podem apresentar antígenos para as células T. As células dendríticas expressam as moléculas coestimuladoras CD80 e CD86 que deflagram a maturação de células T efetoras específicas de antígenos, ligando-se a CD28 na superfície das células T. As células dendríticas se dividem em dois grupos: células dendríticas convencionais (cDCs, do inglês conventional dentritic cells), que residem no tecido linfoide, e pDCs que podem residir no tecido linfoide e migrar para tecidos não linfoides. Observou-se um número bastante elevado de pDCs no sangue periférico de pacientes com artrite reumatoide que haviam sido tratados com metotrexato ou infliximab e que chegaram ao ponto de remissão clínica. A cultura concomitante desses pDCs com células T virgens levou a uma indução do fenótipo Treg.24 Por outro lado, acredita-se que os pDCs sejam patogênicos nos casos de lúpus eritematoso sistêmico devido à capacidade para liberar grandes quantidades de interferons tipo 1.25
A ativação de células T é uma etapa importante na patogênese de artrite reumatoide (AR). A ativação de células T depende da participação do complexo de receptores de células T (TCR, do inglês T cell receptor) e da interação entre as moléculas coestimuladoras nas células T e nas células apresentadoras de antígenos (APCs, do inglês antigen-presenting cells) como as células dendríticas. O abatacept (CTLA-4-Ig) é um modulador de coestimulação que inibe a ativação das células T ligando-se a CD80 e CD86 nas APCs. O abatacept impede competitivamente a ligação de CD28 a uma célula T, evitando a ativação completa desse tipo de célula. A utilidade desse agente nos casos de artrite reumatoide dá suporte a um papel importante das células T nesse tipo de doença.
O rituximab é um anticorpo monoclonal quimérico direcionado contra o antígeno CD20. O antígeno CD20 é encontrado na superfície dos linfócitos B, iniciando no estágio da célula pré-B, embora não esteja presente nas células B que se diferenciam em células plasmáticas. O anticorpo do rituximab é uma imunoglobulina IgG1 kappa e o domínio Fab liga-se a CD20 nos linfócitos B. O rituximab pode provocar a deleção de células B através dos mecanismos descritos acima, porém não se sabe quais mecanismos são dominantes para a depleção de células B em pacientes com artrite reumatoide ou vasculite. O domínio Fc do rituximab possivelmente não recrute células efetoras para promover a lise de linfócitos B através da citotoxicidade celular dependente de anticorpos (CCDA) ou pode ser importante na citotoxicidade mediada por células dependente de complementos. Além disso, a ligação do anticorpo CD20 poderá resultar na interrupção do crescimento ou em apoptose independente da citotoxidade. O rituximab foi aprovado pela FDA para uso no tratamento de doenças como artrite reumatoide, granulomatose com poliangite e poliangite microscópica [ver as Tabelas 3 e 4].
O belimumab é um anticorpo humano monoclonal IgG1 lambda que se liga ao estimulador de linfócitos B humanos (BLyS) inibindo a atividade biológica. O BLyS é uma proteína transmembranar expressa em monócitos, células dendríticas e células estromais da medula óssea. A forma transmembranar pode ser clivada e gerar proteínas solúveis. O BLyS inibe a apoptose de células B e promove a diferenciação de células B em células plasmáticas produtoras de imunoglobulina.26 O BLyS é o único ligante do receptor 3 do BLyS (BR3). A interação entre o BLyS e o BR3 é imprescindível para a sobrevivência de células B virgens e de células B primárias maduras. O BLyS é também o ligante do ativador transmembranar-1 e do modulador de cálcio e do interagente ligante da ciclofilina (TAC1), que é encontrado em um subgrupo de células T e de linfócitos B maduros, e do antígeno para maturação de células B (BCMA) encontrado nas células plasmáticas e em linfócitos B maduros. A interação entre o BLyS e o TAC1 ou o BCMA está envolvida nas ações das células B ativadas por antígenos, das células B da memória e nas células B plasmáticas. O BLyS é o único ligante para o BR3, embora o BLyS e APRIL (um ligante indutor de proliferação) sejam ligantes para o TAC1 e BCMA. Ao usar o BLyS como alvo, a expectativa é que o belimumab poderia afetar as células B logo no início da ontogenia, considerando que as células B de estágio tardio poderiam receber sinais provenientes do ligante APRIL através do TAC1 e BCMA. O belimumab é o primeiro medicamento novo aprovado pela FDA para uso no tratamento de lúpus eritematoso sistêmico nas últimas décadas [ver as Tabelas 3 e 4].
As atividades biológicas atribuídas ao TNF-a incluem indução de outras citocinas pró-inflamatórias, como a IL- e a IL-6, intensificação na migração de leucócitos aumentando a expressão das moléculas de adesão nas células endoteliais e leucócitos, ativação de neutrófilos e eosinófilos, e indução de reagentes da fase aguda. A TNF-a ativa também padrões pró-inflamatórios da expressão gênica em células teciduais como fibroblastos, condrócitos e osteoclastos. Há dois receptores distintos para o TNF-a: p55 e p75 que ligam as moléculas oligoméricas do TNF-a nas superfícies celulares e nas formas solúveis. A atividade biológica do TNF-a depende da ligação com os receptores superficiais celulares p55 ou p75 do TNF [ver as Tabelas 3 e 4].
O infliximab é um anticorpo monoclonal quimérico IgG1 kappa com especificidade para formas solúveis e transmembranares do TNF-a que impedem sua ligação aos receptores superficiais celulares p55 ou p75 do TNF. O infliximab é um anticorpo anti-TNF-a quimérico usado em camundongos e seres humanos produzido a partir de uma região murina variável (porção de ligação a antígenos) e de um domínio Fc da IgG1 humana (região constante). O infliximab foi aprovado pela FDA para o tratamento de condições como artrite reumatoide, espondilite anquilosante, artrite psoriática, psoríase, doença de Crohn e colite ulcerativa TNF [ver as Tabelas 3 e 4].
O adalimumab é um anticorpo IgG1 monoclonal recombinante humano direcionado contra o TNF-a. O adalimumab bloqueia a interação do TNF-a com os receptores superficiais celulares p55 ou p75 do TNF, neutralizando, consequentemente, a atividade do TNF-a. O adalimumab foi aprovado pela FDA para o tratamento de condições como artrite reumatoide, artrite juvenil idiopática, espondilite anquilosante, artrite psoriática, psoríase e doença de Crohn [ver as Tabelas 3 e 4].
O golimumab é um anticorpo IgG1 monoclonal recombinante humano kappa direcionado contra o TNF-a. O golimumab liga-se às formas solúveis e transmembranares do TNF-a. O golimumab foi aprovado pela FDA para o tratamento de condições como artrite reumatoide, artrite psoriática e espondilite anquilosante [ver as Tabelas 3 e 4].
|
Tabela 3: Indicações Aprovadas pela FDA para Agentes Biológicos
| ||||||||
|
Agente Biológico |
Artrite Reumatoide |
Artrite Juvenil Idiopática |
Espondilite Anquilosante |
Artrite Psoriática |
Psoríase |
Doença de Crohn |
Colite Ulcerativa |
LES |
|
Abatacept |
X |
X |
|
|
|
|
|
|
|
Adalimumab |
X |
X |
X |
X |
X |
X |
|
|
|
Anakinra |
X |
|
|
|
|
|
|
|
|
Belimumab |
|
|
|
|
|
|
|
X |
|
Certolizumab |
X |
|
|
|
|
X |
|
|
|
Etanercept |
X |
X |
X |
X |
X |
|
|
|
|
Golimumab |
X |
|
X |
X |
|
|
|
|
|
Infliximab |
X |
|
X |
X |
X |
X |
X |
|
|
Rituximab |
X |
|
|
|
|
|
|
|
|
Tocilizumab |
X |
X |
|
|
|
|
|
|
|
Ustekinumab |
|
|
|
|
X |
|
|
|
FDA =Food and Drug Administration; LES = lúpus eritematoso sistêmico.
O certolizumab é um fragmento Fab’ de anticorpos humanizados recombinantes peguilhados específico para o TNF-a. O certolizumab liga-se às formas solúveis e transmembranares do TNF-a e, consequentemente, inibe de forma seletiva a ligação do TNF-a a seus receptores. A peguilação aumenta a meia-vida do certolizumab e elimina a porção Fc quimérica. Em razão da ausência de uma porção Fc, o certolizumab não induz a ativação de complementos, citotoxicidade celular dependente de anticorpos (CCDA) ou apoptose. O golimumab foi aprovado pela FDA para o tratamento de condições como artrite reumatoide e doença de Crohn [ver as Tabelas 3 e 4].
O etanercept é uma proteína de fusão consistindo de uma porção extracelular de ligação ao ligante do receptor do TNF humano p75 ligada à porção Fc da IgG1 humana. O etanercept é uma forma solúvel dimérica do receptor do TNF p75que pode ligar duas moléculas do TNF. O etanercept inibe a ligação do TNF-a e do TNF-ß (linfotoxina-a) aos receptores do fator de necrose tumoral das células superficiais. O etanercept foi aprovado pela FDA para o tratamento de condições como artrite juvenil idiopática, espondilite anquilosante, artrite psoriática e psoríase [ver as Tabelas 3 e 4].
|
Tabela 43: Mecanismos dos Agentes Biológicos Usados no Tratamento de Doenças Autoimunes
|
| ||
|
Agente |
Construção Molecular |
Mecanismo de Ação |
|
|
Abatacepte |
Proteína de fusão recombinante |
Inibição da ativação de linfócitos pela ligação a CD80/CD86, interação de bloqueio com CD28.
| |
|
Adalimumab |
mAB humano recombinante |
Ligação com o TNF-a neutralizando sua atividade.
| |
|
Anakinra |
Antagonista do receptor humano da IL-6 recombinante
|
Bloqueio da atividade biológica da IL-1 inibindo a ligação da IL-1 ao receptor tipo 1 da IL-1.
| |
|
Belimumab |
mAB humano recombinante |
Liga-se ao estimulador de linfócitos B humanos (BLyS), inibindo a atividade biológica.
| |
|
Certolizumab |
Fragmento Fab’ do anticorpo humanizado recombinante peguilado
|
Liga-se ao TNF-a neutralizando sua atividade.
| |
|
Etanercept |
Proteína de fusão recombinante
|
Liga-se ao TNF-a e à linfotoxina-a neutralizando sua atividade.
| |
|
Golimumab |
mAB humano recombinante |
Liga-se ao TNF-a neutralizando sua atividade.
| |
|
Infliximab |
mAB quimérico |
Liga-se ao TNF-a neutralizando sua atividade.
| |
|
Rituximab |
mAB quimérico |
Liga-se a CD20 nos linfócitos B levando à lise de células B.
| |
|
Tocilizumab |
mAB humano recombinante |
Liga-se ao receptor de IL-6.
| |
|
Ustekinumab |
mAB humano recombinante |
Liga-se à subunidade p40 de IL-12 e IL-23.
| |
IL = interleucina; mAB (monoclonal antibody) = anticorpo monoclonal; TNF (tumor necrosis factor) – fator de necrose tumoral.
O tocilizumab é um anticorpo monoclonal IgG1 kappa humanizado para o receptor de IL-6. O tocilizumab liga as formas solúveis e membranares do receptor de IL-6, impedindo a ligação da IL-6 com seu receptor e bloqueando seus efeitos. A interleucina-6 (IL-6) pode ativar células T, células B, macrófagos e osteoclastos, além de fazer a mediação da resposta hepática da fase aguda aumentando a produção de pentraxinas. A IL-6 é expressa no sinóvio da artrite reumatoide e pode provocar erosões pela ativação de osteoclastos. Os efeitos benéficos na artrite reumatoide incluem redução na atividade da doença e prevenção de erosões ósseas [ver as Tabelas 3 e 4].
O ustekimumab é um anticorpo monoclonal direcionado contra a subunidade p40 das interleucinas-12/23. A IL-12/IL-23 são citocinas que desempenham papeis importantes nas fases iniciais das respostas imunes e podem direcionar a regulação e a diferenciação das células Th1 e Th17, respectivamente. A IL-12 e IL-23 compartilham uma cadeia comum de peptídeos, a subunidade p40, sendo que o ustekimumab tem como alvo a atividade de ambas as citocinas reconhecendo a subunidade p40 compartilhada. O ustekimumab é eficaz no tratamento de psoríase [ver as Tabelas 3 e 4].
O medicamento abatacepte é uma proteína de fusão solúvel usada no tratamento de artrite reumatoide que consiste de CTLA-4 humano ligado a uma porção Fc modificada da IgG1 humana. O abatacepte é um modificador de coestimulação que inibe a ativação de linfócitos T ligando CD80 e CD86 com afinidade mais elevada que CD28, bloqueando, consequentemente, a interação com CD28 nos linfócitos T. Essa ação impede a ativação completa dos linfócitos T por meio do bloqueio do sinal de coestimulação gerado quando o CD80/CD86 interagir com CD28 [ver as Tabelas 3 e 4].
Os glicocorticoides são usados como agentes anti-inflamatórios e imunossupressivos no tratamento de inúmeras doenças reumáticas. Esses mecanismos de ação possivelmente dependam da dosagem. Os glicocorticoides exercem efeitos sobre a expressão gênica em células imunes e seus efeitos não genômicos são rápidos. Os efeitos genômicos são mediados pelo receptor citosólico de glicocorticoides (GCR, do inglês cytosolic glucocorticoid receptor). Após a ligação ao GCR, o receptor citosólico de glicocorticoides se desloca para o núcleo e induz a síntese de proteínas anti-inflamatórias como a lipocortina e o inibidor de NF-kB. Em seguida, essas proteínas anti-inflamatórias alteram a função dos fatores transcritos como ativadoras da proteína-1, do fator-3 regulador do interferon e do NF-kB. Portanto, o GCR glicocorticoide/citosólico pode regular indiretamente a expressão de citocinas pró-inflamatórias como a IL-1, IL-2, IL-6, TNF-a e interferon gama.27,28 Em doses muito elevadas, acima de 30 mg de prednisona todos os dias ou de um medicamento equivalente, os glicocorticoides podem alterar as propriedades das membranas plasmáticas e mitocondriais, alterando, consequentemente, a função e a atividade das proteínas associadas a membranas. Poderá haver uma redução no ciclo do cálcio e do sódio através das membranas plasmáticas e na produção de trifosfato de adenosina. Por exemplo, a dexametasona altera a sinalização de cálcio por meio do enfraquecimento da força do sinal do TCR no sentido do fluxo, o que poderá suprimir a resposta imune.28 Os glicocorticoides podem também inibir a liberação de ácido araquidônico, que é um precursor essencial de muitos mediadores de crescimento celular e inflamação.
A 6-mercaptopurina é um metabólito da azatioprina que inibe a síntese da purina, impedindo, consequentemente, a síntese de DNA e a proliferação de células. A azatioprina reduz o número de linfócitos B e T em circulação (em particular as células CD8+), a síntese da IgM e IgC, a função das células NK e a secreção de IL-2.29,30 A azatioprina foi aprovada pela FDA para uso no tratamento de artrite juvenil idiopática, embora geralmente seja mais utilizada em condições sistêmicas como vasculite.
A ciclofosfamida ainda não está aprovada pela FDA para uso no tratamento de doenças reumáticas, porém é usada para tratar casos graves de LES e vasculite. Os metabólitos ativos da ciclofosfamida entram em ligação cruzada com o DNA, de modo que não podem replicar. A ciclofosfamida é citotóxica para os linfócitos em repouso e em processo de divisão. A ciclofosfamida suprime a função das células Th, diminui o número de células T ativadas e diminui o número de células B.31
A leflunomida é um derivado do isoxazol desenvolvido como um agente modificador de doença utilizado no tratamento de artrite reumatoide. O mecanismo de ação da leflunomida se baseia na inibição da diidroorotato desidrogenase, enzima responsável pela síntese de novo do monofosfato de uridina de ribonucleotídeos de pirimidina. Os linfócitos dependem da síntese de novo da pirimidina para atender suas necessidades metabólicas para expansão clonal e diferenciação em células efetoras. A leflunomida foi aprovada pela FDA para uso no tratamento de artrite reumatoide e foi aprovada também na Europa para uso nos casos de artrite psoriática.
O metotrexato foi desenvolvido como antagonista do ácido fólico para impedir a síntese da purina e da pirimidina, que é vital para a proliferação celular. As propriedades anti-inflamatórias de baixas doses de metotrexato não podem ser explicadas apenas por seus efeitos antiproliferativos. O metotrexato é um inibidor potente da aminoimidazol carboxamida ribonucleotídeo (AICAR, do inglês, aminoimidazole carboxamide ribonucleotide) transformilase. Isso resulta no acúmulo intracelular da AICAR. A AICAR inibe a monofosfato de adenosina (AMP, do inglês adenosine monophosphate) deaminase, causando o acúmulo de AMP que, em seguida, é liberado e convertido em adenosina extracelular. A liberação de adenosina estimulada pelo metotrexato em sítios de inflamação provoca a supressão das funções de neutrófilos, monócitos, macrófagos, células dendríticas e linfócitos.32 O metotrexato foi aprovado pela FDA para uso no tratamento de artrite reumatoide, artrite juvenil idiopática e psoríase.
Ainda não se conhece o mecanismo exato de ação da sulfasalazina. A produção de adenosina em sítios de inflamações possivelmente seja uma propriedade anti-inflamatória que a sulfasalazina compartilha com o metorexato.33 A sulfasalazina inibe também a expressão do TNF-a induzindo a apoptose de macrófagos e suprime a produção de IgM e IgG pelas células B.35,36 A despeito desses possíveis mecanismos, de maneira geral não se considera a sulfasalazina como inibidora das defesas normais de hospedeiros contra patógenos microbianos. A sulfasalazina foi aprovada pela FDA para uso no tratamento de artrite reumatoide, artrite juvenil idiopática e colite ulcerativa.
A ciclosporina forma um complexo de proteínas citoplásmicas com a ciclofilina. A ciclosporina se liga à calcineurina, que é uma fosfatase intracelular. O tacrolimo forma um complexo com FKBP-12b, que interage com a calcineurina. Esses complexos regulam a transcrição do gene da IL-2.37 Consequentemente, a ciclosporina e o tacrolimo bloqueiam funções que dependem da IL-2, incluindo a amplificação de respostas imunes celulares e a geração de células T efetoras. Esses medicamentos inibem a produção indireta de anticorpos pelas células B, reduzindo a disponibilidade funcional das células B auxiliares.38 A ciclosporina foi aprovada pela FDA para o tratamento de artrite reumatoide e psoríase, embora nos dias atuais raramente seja utilizada nos casos de artrite reumatoide. O tacrolimo foi aprovado pela FDA para uso no tratamento de doenças reumáticas e no Canadá foi aprovado para os casos de artrite reumatoide.
O micofenolato mofetil se decompõe em ácido micofenólico (AMF) que é sua forma ativa. O AMF se liga de forma reversível ao monofosfato de inosina desidrogenase, que é uma enzima imprescindível para a síntese de novo dos nucleotídeos de purina em linfócitos, mas não na maior parte dos outros tipos de células. A disponibilidade reduzida dos nucleotídeos de purina inibe a proliferação de células T e B. O micofenolato mofetil se liga com afinidade mais alta a uma isoforma de monofosfato de inosina desidrogenase, que é expressa em linfócitos ativados. Acredita-se que esse medicamento tenha também efeitos no sentido do fluxo, incluindo a expressão de moléculas de adesão nas células endoteliais. Consequentemente, os ligantes das células T correspondentes se alteram sob o ponto de vista funcional. Isso prejudica a adesão das células T ao endotélio, a diapedese através do endotélio e o recrutamento de linfócitos para os sítios de inflamação.39,40 O micofenolato mofetil não foi aprovado pela FDA para uso no tratamento de doenças reumáticas, embora seja utilizado extensivamente nos casos de nefrite lúpica.
A hidroxicloroquina e a cloroquina são agentes antipalúdicos que também são usados no tratamento de doenças autoimunes. Os antipalúdicos – também conhecidos por antimaláricos – são bases fracas que conseguem penetrar nos lisossomas no interior das células, onde se tornam protonados, elevam o pH e, consequentemente, interferem nas funções lisossômicas que dependem de um ambiente ácido. Os antígenos são processados para cargas de peptídeos nas moléculas classe II do complexo de histocompatibilidade principal nos lisossomos ácidos de células dendríticas, células B e monócitos. Portanto, os antipalúdicos possivelmente interfiram na apresentação de antígenos para células T autorreativas inibindo o processamento e a apresentação de antígenos.
Os receptores toll-like (TLRs, do inglês toll-like receptors) intracelulares também são componentes dos lisossomos ácidos e podem reconhecer os componentes do ácido nucleico, levando à patogênese de doenças autoimunes como lúpus eritematoso sistêmico. Por exemplo, o TLR9 é reativo para fragmentos de DNA da citosina-guanina fosfodiéster (CpG).41 Levantou-se a hipótese de que os antipalúdicos inibam a ativação de TLRs intracelulares. A hidroxicloroquina foi aprovada pela FDA para uso no tratamento de artrite reumatoide e lúpus eritematoso sistêmico. A cloroquina não foi aprovada pela FDA para o tratamento de doenças reumáticas. Aparentemente, os antipalúdicos não comprometem as defesas dos hospedeiros contra infecções.
O tratamento de doenças autoimunes está evoluindo na medida em que os mecanismos de patogênese se tornam mais claros nos níveis celular e molecular e levam ao desenvolvimento de terapias específicas. Por outro lado, o uso de agentes biológicos no tratamento de autoimunidades possibilitou a aquisição de conhecimentos mais profundos dessas doenças. Pode-se prever o progresso rápido nessa área da terapêutica clínica, com metas de atingir remissões no longo prazo ou mesmo remissões permanentes e, em última análise, a cura de condições autoimunes.
|
Os autores não mantêm nenhuma relação comercial com os fabricantes dos produtos ou com os fornecedores dos serviços discutidos neste capítulo. |
1.Muench MO, Barcena A. Broad distribution of colonyforming cells with erythroid, myeloid, dendritic cell, and NK cell potential among CD34(++) fetal liver cells. J Immunol 2001;167:4902–9.
2. Kawai T, Akira S. The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat Immunol 2010;11:373–84. 3. Beutler B. Microbe sensing, positive feedback loops, and the pathogenesis of infl ammatory diseases. Immunol Rev 2009; 227:248–63.
4. Schatz DG, Ji Y. Recombination centres and the orchestration of V(D)J recombination. Nat Rev Immunol 2011;11:251– 63.
5. Anthony RM, Nimmerjahn F, Ashline DJ, et al. Recapitulation of IVIG anti-infl ammatory activity with a recombinant IgG Fc. Science 2008;320:373–6.
6. Chen Y, Khanna S, Goodyear CS, et al. Regulation of dendritic cells and macrophages by an anti-apoptotic cell natural antibody that suppresses TLR responses and inhibits infl ammatory arthritis. J Immunol 2009;183:1346–59.
7. Horiuchi T, Mitoma H, Harashima S, et al. Transmembrane TNF-alpha: structure, function and interaction with anti-TNF agents. Rheumatology (Oxford) 2010;49:1215–28.
8. Leandro MJ, Cooper N, Cambridge G, et al. Bone marrow B-lineage cells in patients with rheumatoid arthritis following rituximab therapy. Rheumatology (Oxford) 2007;46:29– 36.
9. Tesmer LA, Lundy SK, Sarkar S, Fox DA. Th17 cells in human disease. Immunol Rev 2008;223:87–113.
10. Wang W, Shao S, Jiao Z, et al. The Th17/Treg imbalance and cytokine environment in peripheral blood of patients with rheumatoid arthritis. Rheumatol Int 2012;32:887–93. Epub 2011 Jan 11.
11. Niu Q, Cai B, Huang ZC, et al. Disturbed Th17/Treg balance in patients with rheumatoid arthritis. Rheumatol Int 2012;32:2731–6. Epub 2011 Aug 2.
12. van Hamburg JP, Asmawidjaja PS, Davelaar N, et al. Th17 cells, but not Th1 cells, from patients with early rheumatoid arthritis are potent inducers of matrix metalloproteinases and proinfl ammatory cytokines upon synovial fi broblast interaction, including autocrine interleukin-17A production. Arthritis Rheum 2011;63:73–83.
13. Kimura A, Kishimoto T. IL-6: regulator of Treg/Th17 balance. Eur J Immunol 2010;40:1830–5.
14. Kryczek I, Bruce AT, Gudjonsson JE, et al. Induction of IL-17+ T cell traffi cking and development by IFN-gamma: mechanism and pathological relevance in psoriasis. J Immunol 2008;181:4733–41.
15. Gierut A, Perlman H, Pope RM. Innate immunity and rheumatoid arthritis. Rheum Dis Clin North Am 2010;36: 271–96.
16. Wijbrandts CA, Vergunst CE, Haringman JJ, et al. Absence of changes in the number of synovial sublining macrophages after ineffective treatment for rheumatoid arthritis: implications for use of synovial sublining macrophages as a biomarker. Arthritis Rheum 2007;56:3869–71.
17. Haringman JJ, Gerlag DM, Zwinderman AH, et al. Synovial tissue macrophages: a sensitive biomarker for response to treatment in patients with rheumatoid arthritis. Ann Rheum Dis 2005;64:834–8.
18. Eissner G, Kolch W, Scheurich P. Ligands working as receptors: reverse signaling by members of the TNF superfamily enhance the plasticity of the immune system. Cytokine Growth Factor Rev 2004;15:353–66.
19. Mitoma H, Horiuchi T, Tsukamoto H, et al. Mechanisms for cytotoxic effects of anti-tumor necrosis factor agents on transmembrane tumor necrosis factor alpha-expressing cells: comparison among infl iximab, etanercept, and adalimumab. Arthritis Rheum 2008;58:1248–57.
20. Arora T, Padaki R, Liu L, et al. Differences in binding and effector functions between classes of TNF antagonists. Cytokine 2009;45:124–31.
21. Nesbitt A, Fossati G, Bergin M, et al. Mechanism of action of certolizumab pegol (CDP870): in vitro comparison with other anti-tumor necrosis factor alpha agents. Infl amm Bowel Dis 2007;13:1323–32.
22. Rosengren S, Hoffman HM, Bugbee W, Boyle DL. Expression and regulation of cryopyrin and related proteins in rheumatoid arthritis synovium. Ann Rheum Dis 2005;64: 708–14.
23. Bartok B, Firestein GS. Fibroblast-like synoviocytes: key effector cells in rheumatoid arthritis. Immunol Rev 2010;233: 233–55.
24. Kavousanaki M, Makrigiannakis A, Boumpas D, Verginis P. Novel role of plasmacytoid dendritic cells in humans: induction of interleukin-10-producing Treg cells by plasmacytoid dendritic cells in patients with rheumatoid arthritis responding to therapy. Arthritis Rheum 2010;62:53–63.
25. Pascual V, Allantaz F, Patel P, et al. How the study of children with rheumatic diseases identifi ed interferon-alpha and interleukin-1 as novel therapeutic targets. Immunol Rev 2008;223:39–59.
26. Do RK, Hatada E, Lee H, et al. Attenuation of apoptosis underlies B lymphocyte stimulator enhancement of humoral immune response. J Exp Med 2000;192:953–64.
27. Strehl C, Spies CM, Buttgereit F. Pharmacodynamics of glucocorticoids. Clin Exp Rheumatol 2011;29(5 Suppl 68): S13–8.
28. Buttgereit F, Burmester GR, Straub RH, et al. Exogenous and endogenous glucocorticoids in rheumatic diseases. Arthritis Rheum 2011;63:1–9.
29. Fox DA, McCune WJ. Immunosuppressive drug therapy of systemic lupus erythematosus. Rheum Dis Clin North Am 1994;20:265–99.
30. Fox DA, McCune WJ. Immunologic and clinical effects of cytotoxic drugs used in the treatment of rheumatoid arthritis and systemic lupus erythematosus. Concepts Immunopathol 1989;7:20–78.
31. Lacki JK, Schochat T, Sobieska M, et al. Immunological studies in patients with rheumatoid arthritis treated with methotrexate or cyclophosphamide. Z Rheumatol 1994;53: 76–82.
32. Cronstein B. How does methotrexate suppress infl ammation? Clin Exp Rheumatol 2010;28(5 Suppl 61):S21–3.
33. Morabito L, Montesinos MC, Schreibman DM, et al. Methotrexate and sulfasalazine promote adenosine release by a mechanism that requires ecto-5’-nucleotidase-mediated conversion of adenine nucleotides. J Clin Invest 1998;101:295– 300.
34. Wahl C, Liptay S, Adler G, Schmid RM. Sulfasalazine: a potent and specifi c inhibitor of nuclear factor kappa B. J Clin Invest 1998;101:1163–74.
35. Hirohata S, Ohshima N, Yanagida T, Aramaki K. Regulation of human B cell function by sulfasalazine and its metabolites. Int Immunopharmacol 2002;2:631–40.
36. Rodenburg RJ, Ganga A, van Lent PL, et al. The antiinfl ammatory drug sulfasalazine inhibits tumor necrosis factor alpha expression in macrophages by inducing apoptosis. Arthritis Rheum 2000;43:1941–50.
37. Tsokos GC. Immunomodulatory treatment in patients with rheumatic diseases: mechanisms of action. Semin Arthritis Rheum 1987;17:24–38.
38. Heidt S, Roelen DL, Eijsink C, et al. Calcineurin inhibitors affect B cell antibody responses indirectly by interfering with T cell help. Clin Exp Immunol 2010;159:199–207.
39. Hochberg MC. Rheumatology. 5th ed. Philadelphia: Mosby/ Elsevier; 2010.
40. Allison AC. Mechanisms of action of mycophenolate mofetil. Lupus 2005;14 Suppl 1:s2–8.
41. Means TK, Latz E, Hayashi F, et al. Human lupus autoantibody-DNA complexes activate DCs through cooperation of CD32 and TLR9. J Clin Invest 2005;115:407–17.
O MedicinaNET é o maior portal médico em português. Reúne recursos indispensáveis e conteúdos de ponta contextualizados à realidade brasileira, sendo a melhor ferramenta de consulta para tomada de decisões rápidas e eficazes.
| Medicinanet Informações de Medicina S/A Cnpj: 11.012.848/0001-57 | info@medicinanet.com.br |
MedicinaNET - Todos os direitos reservados.
Em função da pandemia do Coronavírus informamos que não estaremos prestando atendimento telefônico temporariamente. Permanecemos com suporte aos nossos inscritos através do e-mail info@medicinanet.com.br.