 (Carregando Índice)... (Carregando Índice)... |
Você está em:
Inicial  acp-medicine Infectologia
acp-medicine Infectologia
Última revisão: 04/01/2016
Comentários de assinantes: 0
Mark A. Beilke, MD
Professor de Medicina e Chefe da Divisão de Doenças Infecciosas da Faculdade de Medicina de Wisconsin.
Christy Barrios, pHD
Pós-doutorado, Divisão de Doenças Infecciosas da Faculdade de Medicina de Wisconsin, Milwaukee, WI.
Artigo original: Beilke MA, MD, Barrios C, pHD. Human retroviral infections other than HIV infection. ACP Medicine. 2013.
[The original English language work has been published by DECKER INTELLECTUAL PROPERTIES INC. Hamilton, Ontario, Canada. Copyright © 2015 Decker Intellectual Properties Inc. All Rights Reserved.]
Tradução: Paulo Henrique Machado.
Revisão técnica: Dr. Lucas Santos Zambon.
A origem do termo retrovírus é a ação de uma única enzima que se denomina transcriptase reversa. O fluxo normal de informações genéticas flui do DNA para o RNA e para a proteína; ao inverter esse processo, a transcriptase reversa faz uma cópia de um DNA de fita dupla (DNA complementar [cDNA, do inglês complementary DNA]) do genoma do RNA viral de fita dupla.1 O DNA viral recentemente criado se integra ao genoma da célula hospedeira infectada.
Os retrovírus, além do HIV, estão associados a doenças hematológicas, neurodegenerativas e, possivelmente, autoimunes. A aquisição de retrovírus pode ser exógena ou endógena. Assim como no caso do HIV, a aquisição exógena de retrovírus ocorre através do contato direto com pessoas infectadas ou com seus líquidos corporais; os retrovírus endógenos são transmitidos juntamente com as informações genéticas do hospedeiro contidas nas células germinativas.
Além do HIV, os retrovírus humanos com os papéis mais bem definidos em doenças clínicas são os vírus linfotrópicos da célula T humana tipos I e II (HTLV-I e HTLV-II). O HTLV-I causa linfoma/leucemia de células T do adulto, linfoma não Hodgkin de células T e síndrome neurodegenerativa atípica, designada paraparesia espástica tropical ou mielopatia associada ao HTLV-I (PET/MAH). O HTLV-II causa leucemia de células granulares grandes (CD8+), linfoma cutâneo por CD8+e variante de leucemia das células pilosas de células T grandes;2 esse vírus pode aumentar o risco de incidência de determinadas doenças infecciosas e raramente causa PET/ MAH.
Recentemente, estudos sorológicos realizados na República dos Camarões, na África, detectaram dois vírus intimamente relacionados ao HTLV-I e ao HTLV-II.3,4 Esses vírus, designados HTLV-III e HTLV-IV, ainda não foram associados a alguma doença. No entanto, levando-se em consideração as dificuldades para a realização de estudos em áreas remotas da África, é prematuro tirar quaisquer conclusões.
Os retrovírus humanos endógenos possivelmente desempenhem algum tipo de papel na patogênese das doenças autoimunes.5,6 Embora as replicações de retrovírus endógenos no interior das células de hospedeiro sejam altamente restritas, observa-se a indução replicatória in vitro nas situações em que as células de hospedeiro forem ativadas por algum estímulo químico.7 Entretanto, a ligação entre retrovírus humanos endógenos e doenças clínicas ainda permanece no campo das especulações.
O vírus espumoso humano (VEH) foi investigado extensivamente como causa de doenças. Todavia, as evidências não são suficientes para atribuir algum papel patogênico a esse vírus.8-10
Durante a década de 1990 houve uma grande preocupação em torno da possível identificação de um novo retrovírus em pacientes soronegativos para o HIV-1 que se apresentaram com uma enfermidade semelhante à AIDS. Esses pacientes foram classificados como portadores de linfocitopenia idiopática causada por células T CD4+.11,12 Partículas retrovirais intracisternais humanas foram identificadas nas culturas linfoides desses pacientes;13 no entanto, não foi possível estabelecer um vínculo causal conclusivo, tendo em vista que a infecção retroviral não foi documentada em todos os pacientes afetados.
Comprovadamente, sob a ótica filogenética, os retrovírus humanos estão relacionados aos retrovírus dos primatas do Velho Mundo. O parente viral mais próximo do HTLV-I é o vírus linfotrópico tipo 1 de células de símios, identificado como um agente infeccioso que ocorre naturalmente em muitas espécies de macacos e símios de grande porte do Velho Mundo. Provavelmente, os antecedentes de infecções na população humana ocorreram a partir de eventos transmissivos entre espécies geograficamente distintas; após seu estabelecimento, o vírus seguiu os padrões migratórios das pessoas ao redor do mundo.14 A probabilidade da transmissão do vírus HTLV-I dos primatas para os seres humanos tem suporte de estudos que demonstraram a passagem do vírus da imunodeficiência dos chimpanzés para os seres humanos e sua transformação em HIV.15
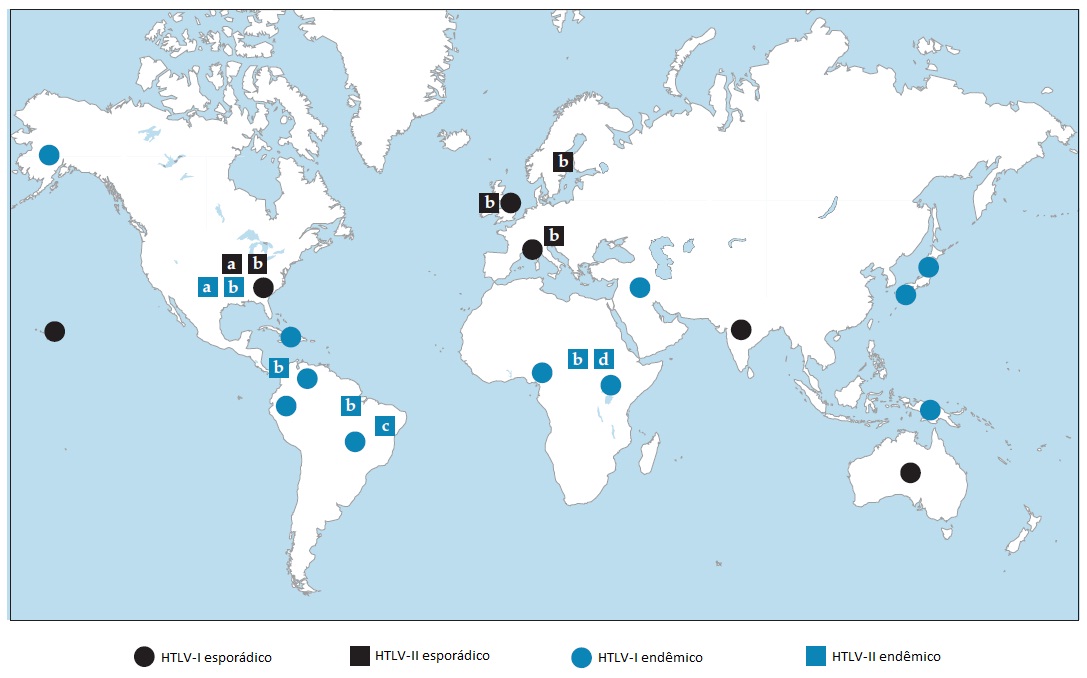
A Figura 1 mostra a distribuição do HTLV-1 em nível mundial. A prevalência varia de 5 a 27%, sendo que as taxas mais elevadas ocorrem em áreas endêmicas, como no Sudoeste do Japão e em Okinawa, onde há mais de 1 milhão de pessoas infectadas. A infecção por HTLV-1 também é prevalente em Taiwan; na bacia do Caribe, incluindo áreas da América do Sul e da região Sudeste dos Estados Unidos; África Central, Israel; e o Círculo Polar Ártico, onde as taxas de soroprevalência variam de 5 a 27% em adultos.16
O HTLV-I é endêmico em várias populações de nativos norte-americanos e em tribos de pigmeus na África Central; nessas populações, as taxas de prevalência variam de 7 a 9%.17 Nos Estados Unidos, a soroprevalência de HTLV-I e HTLV-II varia de 7 a 49% em usuários de drogas injetáveis e em prostitutas.16 Os estudos sorológicos e epidemiológicos iniciais não conseguiram fazer uma distinção eficiente entre HTLV-I e HTLV-II, embora esse problema tenha sido solucionado através do uso de ensaios sorológicos mais específicos.17 Esses ensaios mostraram que a grande maioria das infecções pelo HTLV em usuários de drogas injetáveis é causada pelo HTLV-II.18
Nos Estados Unidos, a soroprevalência de HTLV-I e HTLV-II em doadores de sangue é de 0,025%.16 Estima-se que a taxa anual de soroconversão HTLV-I/II na população seja de 1,09 por 100.000 pessoas.19 Como nos Estados Unidos as doações de sangue são rastreadas para verificar a presença de HTLV-I e HTLV-II, estima-se que o risco de transmissão desse tipo de infecção seja de 1 em 641.000 indivíduos (intervalo de confiança de 95%, de 256.000 a 2.000.000).19 As vias mais comuns de transmissão de infecção por HTLV-I são de mães para filhos durante a gestação, pela via perinatal ou através da amamentação com leite materno, pelo contato sexual e através de agulhas contaminadas.20

Figura 1: Distribuição mundial de infecções causadas pelo vírus linfotrópico de células T humanas tipos I e II (HTVL-I e HTVL-II).113,177-180 A infecção pelo HTVL-I ocorre principalmente em localizações geográficas onde o vírus é endêmico, com ocorrência esporádica de infecções em algum outro local como resultado da imigração ou do uso de drogas injetáveis. Por outro lado, as infecções pelo HTVL-II ocorrem não apenas em populações endógenas, mas se disseminam também em proporções epidêmicas entre usuários de drogas nas grandes áreas metropolitanas das Américas e da Europa. Cabe observar que o HTVL-II possui quatro subtipos moleculares (a-d).
A distribuição de LTA e de PET/MAH se sobrepõe à distribuição de HTVL-I, sendo que mais de 95% das pessoas afetadas apresentam evidências de infecção por HTVL-I. O desenvolvimento de LTA em pessoas infectadas através do uso de produtos derivados do sangue é uma ocorrência muito rara; mesmo assim, 20% de pacientes com PET/MAH adquirem HTVL-I de sangue contaminado.
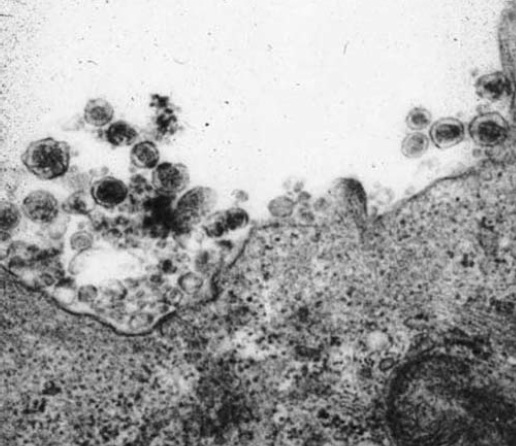
Todos os retrovírus humanos compartilham várias similaridades em relação à estrutura, à organização genômica e ao modo de replicação.21 Sob o ponto de vista estrutural, os retrovírus se classificam nos tipos A, B, C ou D com base na forma e no local onde ocorrem a maturação das partículas no interior da célula do hospedeiro.22-24 Os núcleos dos vírus tipo C (como o HTLV-I e HTLV-II) se caracterizam por duas membranas concêntricas com aparência de anel duplo nos espécimes preparados para microscopia eletrônica25 [ver a Figura 2]. Por outro lado, os retrovírus endógenos, que são partículas do tipo A, são menores (70 mm) e apenas intracelulares.26 Os vírus tipo C montam suas cápsides e amadurecem na membrana plasmática das células infectadas durante o processo de germinação. Não se observa a presença de nenhum precursor intracelular.27
Figura 2: Imagem por micrografia eletrônica de vírions de vírus linfotrópico de células T humanas tipo I (HTVL-I) germinando a partir de culturas de linfócitos obtidas de um paciente infectado pelo HTVL-I. Os vírions do HTVL-I possuem a característica nucleocapsídea redonda central dos retrovírus tipo C. Os vírions do HTVL-I nunca são identificados em células não cultivadas ou nos tecidos de indivíduos infectados pelo HTVL-I.
O HTLV-I e HTLV-II diferem do HIV-1 no que diz respeito à morfologia e à estrutura genômica [ver a Tabela 1].28,29 As partículas do vírus HTLV têm um diâmetro aproximado de 100 mm e um invólucro lipídico que inclui a membrana das células plasmáticas do hospedeiro.30 O invólucro lipídico circunda o núcleo capsídeo que contém duas cópias do genoma do RNA de fita dupla.
O genoma retroviral contém sequências não codificadoras e codificadoras. As sequências não codificadoras – sítios importantes de reconhecimento para síntese, integração e poliadenilação de DNA ou RNA – se localizam nas extremidades terminais 5’ e 3’ do genoma. Todos os retrovírus são terminalmente redundantes e possuem sequências idênticas que se denominam repetições terminais longas (LTRs, do inglês long terminal repeats). As sequências codificadoras incluem o gene gag, que codifica antígenos estruturais específicos de grupos; o gene pol, que codifica a polimerase ou transcriptase reversa, integrase e protease de DNA dependente de RNA; e o gene env, que codifica as proteínas estruturais do invólucro. O gene gag codifica um polipeptídeo precursor cuja segmentação mitótica é feita pela protease do vírus codificado para formar diversas proteínas estruturais internas – isto é, proteína de matriz, proteína capsídea e proteína de ligação com o ácido nucleico.21
|
Tabela 1: Características Morfológicas e Estruturais do HIV-1 em Comparação com o HLTV-I e o HLTV-II27,28 | |||
|
Característica
|
HIV-1 |
HLTV-I/HLTV-II | |
|
Taxonomia |
Lentivírus |
Oncornavírus
| |
|
Morfologia |
Nucleocapsídeo em forma de bala com diâmetro variando de 100 a 120 mm.
|
Nucleocapsídeo redondo com diâmetro de 100 mm. | |
|
Vírions detectados pela microscopia eletrônica em amostras de biópsia
|
Sim |
Não | |
|
Genes estruturais (produtos) |
gag (p17, p24, p2, p7, p1, p6)
pol (transcriptase reversa, integrasse, protease)
env (gp120, gp41) |
gag (p19, p24, p15)
pro (protease)
pol (transcriptase reversa, possivelmente integrasse)
env (gp46, gp21)
| |
|
Genes acessórios (produtos e funções) |
vif (proteína de infectividade vírica)
vpr (proteína R viral; melhora a replicação viral).
tat (transativador viral)
rev (regulador da expressão de proteínas virais).
vpu (proteína U viral; melhora a liberação de vírions das células)
nef (fator regulador negativo).
|
tax (ativador transcripcional)
rex (modificação pós-transcripcional)
Possivelmente existam outros genes acessórios, porém suas funções são desconhecidas. | |
|
HTLV (human T-cell lymphotropic vírus) = vírus linfotrópico de células T humanas |
| ||
O genoma do HTLV é altamente conservado, porém a maior convergência de nucleotídeos nas repetições terminais longas possibilitou o desenvolvimento de polimorfismos no comprimento dos fragmentos de restrição (RFLP, do inglês restriction fragment length polymorphisms). Os testes de RFLP permitiram que os pesquisadores classificassem o HTLV-I e o HTLV-II em subtipos genotípicos, o que acabou gerando informações valiosas sobre a transmissão viral.31 Há cinco subtipos moleculares e geográficos principais de HTLV-I: cosmopolita (em todo o mundo), japonês, do Oeste africano, centro africano e melanésio.32 Há três subtipos de HTLV-II que refletem grupos populacionais ao invés de grupos geográficos: a, b e c. O subtipo a é encontrado em usuários de drogas injetáveis em todo o mundo, o subtipo b é encontrado principalmente em índios nativos norte-americanos e o subtipo c é encontrado em tribos indígenas brasileiras.33
O HTLV-I e HTLV-II possuem genomas semelhantes, sendo que ocorre a mesma coisa em aproximadamente 60% dos respectivos nucleotídeos. O gene gag codifica as proteínas p19, p24 e p15. O gene pol codifica a protease e a transcriptase reversa. O gene env codifica o invólucro externo e as glicoproteínas transmembrânicas gp46 e gp21. As proteínas regulatórias removidas (spliced) tax e rex e as outras três fases de leitura aberta constituem o genoma 9kb. As proteínas gag e env são basicamente imunogênicas; os anticorpos a essas proteínas geralmente são detectados por imunoensaio enzimático (EIA, do inglês enzyme immunoassay) e pelo ensaio Western blot.
A transmissão de retrovírus para os seres humanos se caracteriza pela exposição parenteral ou íntima das membranas mucosas ao vírus. A transmissão por fômites é improvável levando-se em conta que, logo após a saída do corpo humano, o invólucro lipídico dos retrovírus é facilmente danificado pela exposição a temperaturas elevadas, detergentes, desinfetantes químicos e através da secagem. Ao contrário do HIV-1, os vírus HTLV são altamente associados às células. Portanto, a transmissão através do contato com líquidos sem células, como o plasma, por exemplo, ocorre em situações extremamente raras. O HTLV-I e HTLV-II (assim como o HIV-1) são transmitidos através dos líquidos genitais; através do sangue (por meio de transfusões, transplantes de órgãos ou por agulhas contaminadas); ou de mães para os filhos através do leite materno, em que há uma quantidade abundante de células T CD4+ infectadas pelo HTLV-I. Existem relatos de casos raros de transmissão in utero.
Preferencialmente, o HTLV-I infecta as células T, principalmente as células T CD4+. Aparentemente, o HTLV-II tem preferência pelas células T CD8+, embora o cDNA viral integrado também possa ser encontrado nas células T CD4+.35,36 Entretanto, a entrada de HTLV-I e HTLV-II no interior das células T CD8+e T CD4+não depende do receptor de CD4 ou CD8. O HTLV-I e o HTLV-II compartilham o tropismo para uma variedade de células não linfoides, sugerindo a possibilidade de que esses vírus utilizem um receptor celular mais amplamente distribuído para ligação e penetração na célula de hospedeiro.37,38 Pelo menos um laboratório apresentou evidências sugerindo que as linhas celulares de quase todos os mamíferos expressam um receptor funcional do invólucro do HTLV; um fato curioso é que o receptor pode ser subexpresso (ou talvez esteja ausente) nas células T de humanos em repouso e seja expresso somente após a ativação das células T.39 O HTLV-I e o HTLV-II podem penetrar nas células T por meio de complexos de receptores diferentes;40 entretanto, as entradas mediadas por receptores podem ser desnecessárias para a infecção das células por esses vírus. O HTLV-I pode se disseminar entre os linfócitos, sem germinar, a partir da célula doadora.41,42 Em vez disso, o vírus poderá ser transmitido através do contato entre as células.
Excetuando-se os casos de transmissão através do sangue, as infecções iniciais por HTLV-I ou HTLV-II são locais, no interior dos linfáticos regionais. O trânsito de células infectadas pelo HTLV-I através dos vasos linfáticos e da corrente sanguínea possivelmente resulte na disseminação do vírus para vários reservatórios de infecção, tais como pele, timo, fígado, baço, tecidos linfoides associados a mucosas e locais perivasculares no interior do sistema nervoso central e periférico.
A replicação eficiente do HTLV-I e do HTLV-II no interior das respectivas células-alvo segue o paradigma da retrovirologia dos mamíferos.20 Após a penetração do vírus na célula T, o RNA viral de fita única é copiado no híbrido RNA/DNA por meio da enzima transcriptase reversa viral. A seguir, após a degradação do modelo de RNA, a transcriptase reversa copia o DNA de fita única no interior de um híbrido RNA/DNA em diversas etapas. Na sequência, o DNA completo de fita dupla circula e se manifesta em uma forma epissomal não integrado ou é transportado para o interior do núcleo da célula e se torna integrado dentro do genoma do hospedeiro em sítios aleatórios.43
Uma das características que define os retrovírus é o cDNA viral de transcrição reversa no interior do genoma da célula de hospedeiro. O cDNA se integra permanentemente em um único sítio aleatório no cromossomo celular. A integração do cDNA viral linear ocorre durante a divisão da célula; consequentemente, em geral, as células não individualizantes são resistentes às infecções retrovirais.
O cDNA viral integrado denomina-se provírus e serve de modelo para a replicação viral. Esse processo de integração poderá resultar em infecção persistente ou em uma transformação maligna da célula infectada. Embora muitos retrovírus possam causar citopatologia na célula de hospedeiro, a replicação da maior parte desses vírus ocorre sem a morte da célula infectada. No caso dos retrovírus humanos endógenos (retrovírus humanos endógenos tipo K [HERV-K], HERV-W e vários outros), diversas cópias de sequências de DNA provirais endógenas são integradas no DNA cromossômico e são transmitidas na linha germinativa. É interessante observar que as sequências provirais endógenas representam 0,1% ou mais das sequências do DNA humano e que, aparentemente, foram adquiridas em algum momento em nosso passado evolucionário.21
A integração dos retrovírus humanos é diferente da integração de determinados retrovírus animais no sentido em que os genes transformadores (oncogenes) estão associados ao genoma viral e o genoma proviral não é inserido regularmente nas proximidades de um gene transformador de hospedeiro. Além disso, os retrovírus humanos clinicamente importantes são exógenos e não são transmitidos em células germinativas, da mesma forma como alguns retrovírus endógenos de vertebrados e os HERVs sem nenhuma importância clínica.
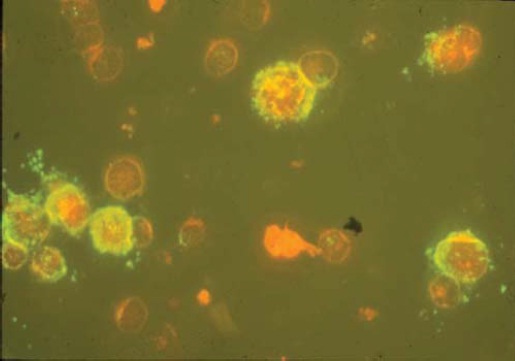
Ao contrário do HIV, que é citopático para as células T CD4+, o HTLV-I e o HTLV-II imortalizam in vitro principalmente as células T do sangue periférico humano. Define-se imortalização como o crescimento de culturas de longo prazo dependente da interleucina-2 (IL-2) [ver a Figura 3]. Subsequentemente, as células se transformam completamente e podem proliferar de forma contínua na ausência da interleucina-2. A transformação celular é uma etapa inicial do processo patogênico do HTLV, o que a distingue de oncogênese ou de malignidade.21 A transformação do HTVL das células T resulta em um pool de células proliferativas não oncogênicas e cria uma população a partir da qual poderá surgir um clone maligno subsequente.21 Esse processo é responsável pelo longo período de latência entre a infecção e o desenvolvimento tumoral em doenças causadas por HTVL.
Figura 3: Micrografia por imunofluorescência de um vírus linfotrópico de células T humanas tipo I dependente da interleucina 2 (HTLV-I) produzindo uma linha celular estabelecida a partir de células mononucleares do sangue periférico de um paciente com mielopatia associada a paraparesia espástica tropical/HTLV-I. A coloração verde representa a expressão do antígeno nuclear p19 do HTLV-I; as células recebem coloração de contraste com azul de Evans e aparecem na cor laranja.
Após a integração do genoma retroviral no DNA de hospedeiro, ocorre o início de uma segunda fase de replicação com a produção de RNA genômico viral, RNA mensageiro (mRNA, do inglês messenger RNA) e síntese proteica. A produção de RNA mensageiro e da síntese proteica ocorre quase que exclusivamente através do mecanismo enzimático da célula de hospedeiro que influencia os produtos regulatórios da genética viral. O processamento de proteínas com partículas virais completas inicia no retículo endoplásmico e no complexo de Golgi. A montagem da estrutura de vírions inicia na membrana plasmática, sendo que os vírions nascentes são liberados a partir das superfícies celulares através do processo de germinação. O invólucro viral de germinação, que possui uma composição fosfolipídea diferente da composição da membrana plasmática da célula, poderá incorporar algumas proteínas da superfície da membrana celular, juntamente com glicoproteínas específicas de vírus.30,44
A complexidade dos retrovírus humanos é mais bem exemplificada pelo conjunto de proteínas virais responsáveis pela regulação da replicação viral e pela resposta da célula de hospedeiro à infecção.21,45 O HTVL-I possui uma região entre o gene env e a LTR 3’que codifica as proteínas regulatórias tax e rex, que são produzidas a partir de mensagens que se juntam de forma diferente a partir de fases de leitura sobrepostas distintas. A proteína tax induz a expressão de fatores de transcrição celular que alteram a expressão genética da célula de hospedeiro, e a proteína rex regula a expressão do mRNA viral.
A proteína tax foi considerada como a proteína transformadora do HTVL-I, além de possuir um envolvimento central nas etapas complexas da leucogênese depois de vários anos de infecção subclínica. Embora outros retrovírus transformadores contenham oncogenes como o src (vírus do sarcoma de Rous), myb (vírus da mieloblastose aviária) e erb-A / erb-B (vírus da eritroblastose aviária), que possui um componente celular ou se integra no genoma de hospedeiro e suprarregula o fluxo normal dos proto-oncogenes e de outros genes promotores do crescimento, aparentemente o HTVL-I não segue nenhum mecanismo de transformação.45 A proteína tax tem uma multiplicidade de funções, incluindo ativação constitutiva dos fatores de transcrição, modificação das vias de transdução de sinal, alteração nas funções das proteínas supressoras de tumores, modulação das proteínas de verificação dos pontos do ciclo celular, interferência nos reparos de DNA e inibição da apoptose. A proteína tax induz a expressão de uma ampla gama de proteínas de células de hospedeiro, incluindo fatores de transcrição e citocinas (p.ex., interleucina 2 [IL-2] e fator de necrose tumoral [TNF]) [ver a Tabela 2].
|
Tabela 2: Genes Celulares Principais Transativados pela Proteína Tax do HTLV-I
| ||
|
Produtos Genéticos |
Efeito Sobre a Atividade
|
Consequência Patofisiológica |
|
Citocinas e receptores
| ||
|
IL-1 |
|
|
|
IL-2 e IL-2Ra |
|
|
|
GM-CSF |
|
|
|
IL-6 |
|
|
|
IL-8 |
Suprarregulação |
Expressão policlonal de células infectadas |
|
IL-10 |
|
|
|
IL-13 |
|
|
|
IL-15 |
|
|
|
TNF-ß |
|
|
|
MIP-1a; MIP-1ß |
|
|
|
Fatores de transcrição de hospedeiro (c-fos c-sis, c-rel, c-myc, erg-1, erg-2, fra-1)
|
Suprarregulação |
Transformação celular |
|
Moléculas de adesão (ICAM-1, vimentina, CD54) |
Suprarregulação |
Rastreamento de linfócitos nos tecidos envolvidos
|
|
Fatores proapoptóticos (bax, p53) |
Infrarregulação |
Transformação celular e leucomogênese
|
|
Enzimas de reparo de DNA |
Infrarregulação |
Transformação celular e leucomogênese
|
GM-CSF (granulocyte-macrophage colony-stimulating fator) = fator estimulador das colônias de granulócitos-macrófagos; HTLV-I (human T-cell lymphotropic vírus type I) = vírus linfotrópico de células T humanas tipo I; ICAM (intercellular adhesion molecules) = moléculas de adesão intercelular; IL = interleucina; IL-2R (interleukin-2 receptor) = receptor da interleucina-2; MIP (major intrinsic protein) = proteínas intrínsecas principais; TNF (tumor necrosis fator) = fator de necrose tumoral.
Entretanto, embora não se conheça como essa indução de fatores de células de hospedeiro possa produzir transformação neoplásica, não há relatos de nenhuma anormalidade cromossômica consistente em casos de linfoma/leucemia de células T do adulto. As observações discordantes entre os meios in vitro e in vivo são temas recorrentes na compreensão da biologia complexa das doenças retrovirais humanas.
Tipicamente, as infecções causadas por HTLV-I e HTLV-II sugerem respostas robustas humorais e celulares de hospedeiros.46 Não obstante, esses vírus são capazes de evitar a destruição por meio de sua capacidade de integração em um genoma de hospedeiro e de propagação durante o processo fisiológico normal de divisão celular, assim como por sua capacidade de disseminação de célula para célula, sem a produção de vírions extracelulares.42,46
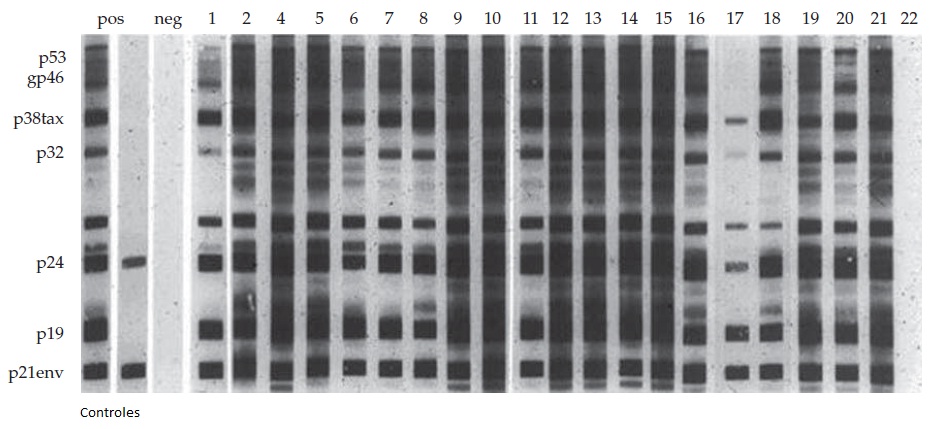
As infecções causadas pelo HTLV-I resultam no desenvolvimento de anticorpos dentro de 1-2 meses, sendo que após esse período de tempo os níveis de anticorpos permanecem estáveis por toda a vida.47 As sequências provirais de ácido nucleico podem ser detectadas durante a janela antes da soroconversão. As manifestações clínicas de doenças se correlacionam intimamente com os títulos dos anticorpos e com a carga proviral.48 Aparentemente, os pacientes que desenvolvem PET/MAH constroem respostas imunes humorais mais robustas [ver a Figura 4], seguidos pelos pacientes com pré-linfoma/leucemia de células T do adulto; de maneira geral, os portadores assintomáticos apresentam titulações mais baixas de anticorpos e níveis mais baixos de carga proviral de HTLV.49,50 Os anticorpos de neutralização viral direcionados para os epítopos no interior da região do invólucro podem ser detectados em portadores assintomáticos do HTLV-I, porém, aparentemente, esses anticorpos não produzem nenhuma proteção.49,50 Todavia, ainda se encontra em fase de investigação o potencial de proteção contra infecções por meio da indução de anticorpos de neutralização viral através de vacinas de HTLV-I.51,52
As infecções causadas pelo HTLV-I e pelo HTLV-II estão associadas a respostas proliferativas espontâneas in vitro, que poderão ser observadas na fase inicial do curso das infecções.34,53 As linhas de células T CD4+ infectadas pelo HTLV-I induzem a proliferação de células T CD4+.53 Há sugestões de que essa indução da proliferação de células T CD4+ seja um mecanismo para a disseminação de infecções pelo HTLV-I, tendo em vista que as células T CD4+recentemente recrutadas poderão se transformar em alvos adicionais para o vírus HTLV-I.34
As células T CD4+específicas para o HTLV-I respondem in vitro aos alvos infectados pelo HTLV-I, sendo que, aparentemente, essas respostas são mais robustas em pacientes com PET/MAH.34 As respostas imunes citotóxicas das células aparentam ser fortes e persistentes em pessoas infectadas pelo HTLV-I.54 Os linfócitos T citotóxicos (LTCs) específicos do HTLV-I controlam a carga viral do HTLV através da eliminação das células T CD4+infectadas pelo HTLV-I. A despeito da alta frequência documentada de linfócitos T citotóxicos específicos do HTLV-I, os LTCs são ineficazes para erradicar todo o reservatório viral. Na realidade, aparentemente, a atividade dos LTCs contribui diretamente para o desenvolvimento de doenças, em especial a PET/MAH.55,56 A expansão dos clones de LTC específicos para a tax de HTLV-I é patognomônica de PET/MAH e se restringe acentuadamente aos alelos específicos do antígeno leucocitário humano (HLA, do inglês human leukocyte antigen).57 Na realidade, o status do HLA de uma pessoa infectada pelo HTLV-I possivelmente seja um fator importante para determinar se a infecção resultará em LTA ou PET/MAH; em um estudo, os alelos específicos do antígeno leucocitário humano (i.e., HLA-A*26, B*4002 e B*4801) foram encontrados em frequências significativamente mais elevadas em pacientes com LTA, em comparação com pacientes com PET/MAH e portadores assintomáticos de HTVL-1.58
Aparentemente, ao contrário dos pacientes com PET/MAH, os pacientes com LTA ou com LTA latente apresentam respostas reduzidas à LTA específicas de HTLV-I.59 Em parte, esse fato se explica pela infrarregulação da expressão do gene tax do HTLV-I nas células LTA ou pela mutação do gene tax.60,61 Uma prova conceitual dessa hipótese foi demonstrada em um modelo de rato com LTA, em que as células T transferidas por adoção de doadores imunizados com a vacina DNA-tax renovaram as respostas da LTA específicas de HTLV-I tax e evitaram o desenvolvimento de LTA.62
Figura 4: Western blots de pacientes colombianos com paraparesia espástica tropical/vírus linfotrópico de células T humanas tipo I (HTVL-I) associada à mielopatia e às respectivas esposas. Observa-se uma resposta imune humoral robusta com reatividade sérica a todos os polipeptídeos principais do HTVL-1. O paciente número 22 é negativo
Assim como os outros vírus, os retrovírus podem produzir infecções latentes, crônicas ou persistentes.63 As infecções latentes se caracterizam pela presença de episódios intermitentes de doença aguda ou subclínica; entre os episódios não é possível detectar nenhum tipo de vírus. Embora, usualmente, nas infecções crônicas o vírus seja demonstrável, não há sintomas visíveis da doença. As infecções persistentes se caracterizam por longos períodos de incubação com crescimento lento das quantidades de vírus, levando, finalmente, a uma doença sintomática. As doenças clínicas se desenvolvem apenas em 5 a 10% de pessoas infectadas pelo HTLV-I ou pelo HTLV-II. Nos casos de LTA, o período de latência entre a infecção e o surgimento de uma doença é de 20 a 30 anos ou mais. O período mediano de latência da PET/MAH é de aproximadamente três anos, embora o período de latência seja de 20 a 30 anos. As manifestações clínicas das infecções por HTLV incluem malignidades, transtornos neurológicos, distúrbios autoimunes e aumento na suscetibilidade a outras doenças infecciosas [ver a Tabela 3].
|
Tabela 3: Manifestações Clínicas das Infecções por HTLV-I e HTLV-II
| ||
|
Manifestação
|
HTLT-I |
HTLV-II |
|
Malignidades |
LTA crônica |
Leucemia de células granulares grandes (CD8+)
|
|
|
LTA latente |
Linfoma cutâneo de célula T CD8+
|
|
|
Linfoma de células T não-Hodgkin |
Leucemia de células pilosas variantes de células T
|
|
Manifestações neurológicas |
PET/MAH |
PET/MAH
|
|
|
|
Neuropatia periférica
|
|
Distúrbios autoimunes |
Polimiosite |
Tireoidite de Hashimoto
|
|
|
Artrite
|
|
|
|
Uveíte
|
|
|
|
Tireoidite
|
|
|
|
Pneumonite |
|
|
Doenças infecciosas |
Infecções oportunistas em pacientes com LTA*
|
Pneumonia bacteriana |
|
|
Dermatite infecciosa em crianças
|
Tuberculose |
|
|
Infecções por Strongyloides stercoralis
|
Infecções no trato urinário |
LTA = leucemia/linfoma de células T de adultos; HTLV (human T-cell lymphotropic vírus) = vírus linfotrópico de células T humanas; PET/MAH = paraparesia espástica tropical ou mielopatia associada ao HTLV-I.
*As infecções oportunistas podem incluir pneumonite por citomegalovírus, herpes zoster disseminado, aspergilose, criptocococitose, infecção disseminada por Mycobacterium atrium e tuberculose miliar.
O vírus linfotrópico de células T humanas tipo I (HTLV-I) é o agente etiológico da leucemia/linfoma de células T de adultos (LTA) e da paraparesia espástica tropical ou mielopatia associada ao HTLV-I (PET/MAH). Esse tipo de vírus tem também implicações em vários outros distúrbios, incluindo artropatia inflamatória, uveíte, polimiosite, doença de Sjögren, distúrbios pulmonares e dermatite infecciosa em crianças. Há associações provocativas – porém não comprovadas – do HTLV-I com micose fungoide e com a síndrome de Sézari. O foco desta discussão é a LTA e a MAH por causa do vínculo etiológico estabelecido com o HTLV-I.
A base fisiopatológica de doenças causadas pelo HTLV-I é bem menos conhecida do que a das doenças causadas pelo HIV-1. Ao contrário do HIV-1, o receptor celular do HTVL-I ainda não foi identificado. Embora as células T CD4+ sejam infectadas produtivamente pelo HTVL-I, ocasionalmente alguns outros tipos de células, tais como as células B e as células T CD8+, também são infectados. Assim como nos casos do HIV-1, o cDNA viral de transcrição reversa se integra, de forma aleatória, à célula de hospedeiro para estabelecer o provírus, porém, ao contrário do HIV-1, o HTVL-I estabelece uma infecção latente com expressão infrequente de produtos genéticos virais. Consequentemente, o HTVL-I tem um nível muito baixo de penetração de doenças; a transformação de uma célula infectada é um evento raro, sendo que o risco acumulado durante a vida de desenvolver LTA varia de apenas 1 a 5% em pessoas infectadas pelo HTVL-I. Estima-se que o período de latência desde a infecção até a doença clínica seja de 30 a 50 anos;64 a maioria das pessoas com LTA aparenta ter adquirido a infecção na infância.65
Há descrições de quatro apresentações clínicas de LTA: aguda, linfomatosa, crônica e latente.64 Todas essas malignidades se caracterizam pela expansão monoclonal de células T CD4+ que contêm o provírus HTLV-I e rearranjos de genes do receptor do padrão clonal de células T. Os vírions do HTLV-I não são recuperados diretamente das células T infectadas, embora a proteína tax viral seja expressa in vivo. Ainda não foi completamente esclarecida a patogênese molecular desse neoplasma induzido por retrovírus.
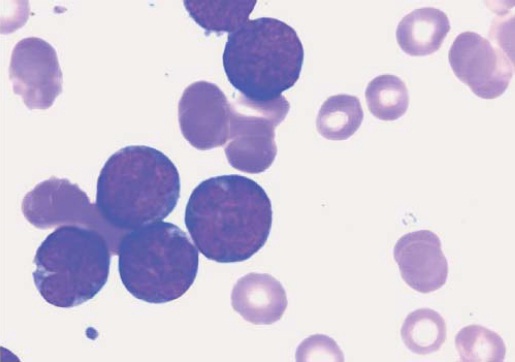
Figura 5: Esfregaço no sangue periférico em leucemia/linfoma de células T de adultos (LTA). As células da LTA (às vezes descritas como células na forma de trevo ou de flor) têm a aparência pleomórfica e multilobular, com convoluções nucleares significativas.
LTA Aguda. A LTA aguda é responsável por aproximadamente 60 a 80% de todos os casos de leucemia/linfoma de células T de adultos. Os pacientes apresentam um pródromo clínico curto, com uma média de duas semanas entre o início dos sintomas e o diagnóstico. A LTA possui uma história natural agressiva; a sobrevida mediana é de seis meses. O quadro clínico se caracteriza pela presença de lesões cutâneas com progressão rápida (essas lesões ocorrem em 40% de casos e variam desde erupções macropapulares a lesões tumorais), infiltrados pulmonares, diarreia, hipercalcemia (observada em 50% de casos), níveis elevados de lactato desidrogenase (DHL) e de fosfatase alcalina, e linfocitose com células mononucleares pleomórficas (denominadas células em forma de trevo ou de flor) que possuem núcleos fendidos e lobulados em forma de cascos de animais [ver a Figura 5]. Talvez seja muito difícil fazer a distinção entre lesões cutâneas e micose fungoide e a síndrome de Sézari [ver a Figura 6]. As lesões pulmonares possivelmente sejam resultado de algum infiltrado leucêmico ou de uma infecção oportunista, como as infecções causadas pelo Pneumocystis jiroveci ou por fungos. A diarreia quase sempre está associada a uma infecção oportunista. Hepatoesplenomegalia é uma presença provável. As lesões ósseas líticas são comuns; essas lesões são irregulares e se compõem de células osteolíticas sem atividade osteoblástica. O envolvimento leptomeníngeo, acompanhado de fraqueza, alteração no estado mental, parestesias e cefaleia ocorre em aproximadamente 10% dos pacientes. Usualmente, o nível proteico do líquido cerebrospinal (LCS) é normal. O diagnóstico é confirmado pela descoberta de células malignas no LCS.
LTA Linfomatosa. A LTA linfomatosa é responsável por aproximadamente 20% de casos de leucemia/linfoma de células T de adultos. A LTA linfomatosa se assemelha à LTA aguda, exceto pela presença de linfadenopatia e pela ausência de linfócitos morfologicamente anormais na circulação. A suspeita diagnóstica pode se fundamentar no local geográfico de nascimento do paciente e na presença de lesões cutâneas e de hipercalcemia. O diagnóstico é confirmado por testes sorológicos do HTLV-I ou pela detecção de provírus integrados no teste da reação em cadeia da polimerase (PCR, do inglês polymerase chain reaction).
LTA Crônica. A LTA crônica é responsável por aproximadamente 15% de casos de leucemia/linfoma de células T de adultos. Esta forma de LTA se caracteriza por níveis séricos normais de cálcio, níveis elevados de lactato desidrogenase (DHL) e pela ausência de envolvimento ósseo, de envolvimento do sistema nervoso central (SNC) ou do trato gastrointestinal (TGI). A sobrevida mediana para esses pacientes é de dois anos. A LTA crônica pode progredir para LTA aguda.
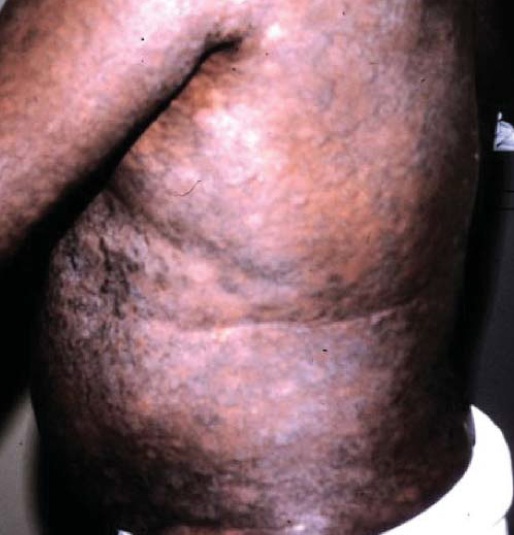
Figura 6: Envolvimento cutâneo em leucemia/linfoma de células T de adultos (LTA). As lesões são generalizadas, nodulares e endurecidas.
LTA Latente. Na LTA latente, que é responsável por menos de 5% de casos de leucemia/linfoma de células T de adultos, uma quantidade inferior a 5% de linfócitos em circulação apresentam anormalidades típicas de LTA ou possuem provírus integrado de HTLV-I, sendo que os pacientes não se apresentam com hipercalcemia, adenopatia ou hepatoesplenomegalia. Não há envolvimento do sistema nervoso central, dos ossos e do trato gastrointestinal, embora seja comum a presença de lesões cutâneas típicas e de infiltrados pulmonares. A sobrevida mediana para pacientes com LTA latente é de cinco anos ou mais.
Ainda persistem os debates sobre o papel desempenhado pela retrovirologia em pacientes com linfoma cutâneo de células T (LCCT) soronegativo para HTLV-I/II. A sugestão ainda não confirmada do envolvimento dos retrovírus no desenvolvimento de casos de LCCT soronegativos para HTLV-I se fundamenta nas evidências discutíveis apresentadas por estudos de pequeno porte.66,67
A paraparesia espástica tropical ou mielopatia associada ao HTLV-I (PET/MAH) é uma mielopatia torácica de progressão lenta associada à paraparesia espástica, perturbações esfincterianas e graus variáveis de disfunção proprioceptiva e sensorial. Um terço dos pacientes se torna enfermo e acamado dentro de dez anos após o diagnóstico.68 A PET/MAH afeta desproporcionalmente as mulheres por razões que ainda não são conhecidas.
De maneira geral, os pacientes com mielopatia associada ao HTLV-I (MAH) apresentam níveis provirais elevados no sangue e respostas imunes mais fortes ao HTLV-I, com titulações mais altas de anticorpos no líquido cerebrospinal do que no soro. Alguns especialistas defendem a teoria de que o envolvimento do SNC nos casos de MAH é o resultado da destruição autoimune de tecidos neuronais pelas células T CD8+ específicas do vírus. O HTLV-I também foi associado a condições como uveíte, artropatia e dermatite infecciosa; provavelmente cada uma dessas condições tenha uma patogênese autoimune ou inflamatória.69
Sob o ponto vista patofisiológico, a PET/MAH é um processo inflamatório progressivo, com infiltração parenquimatosa de células mononucleares no interior da massa cinzenta e branca do SNC, resultando na degeneração simétrica grave da massa branca das colunas laterais, dos tratos corticoespinais e das colunas posteriores.70 Uma das características mais marcantes desse processo são as alterações inflamatórias crônicas na medula espinhal, principalmente a menigomielite da medula torácica inferior. A presença de um infiltrado inflamatório nas meninges espinhais é comum e o parênquima da medula espinhal apresenta evidências de destruição da mielina, assim como ocorre nos casos de esclerose múltipla. Na fase inicial da doença, as alterações inflamatórias são evidentes principalmente ao redor de vênulas e capilares. Ao final, essas alterações inflamatórias perivasculares são acompanhadas de hialinose nos vasos sanguíneos, fibrose meníngea e cicatrizes gliais.
O início das manifestações clínicas nos casos de PET/MAH é insidioso. Os sintomas incluem fraqueza em uma ou em ambas as pernas, dor lombar e incontinência urinária. De maneira geral, a neuropatia periférica é branda. As descobertas físicas incluem paraparesia espástica ou paraplegia com hiperreflexia, clônus no tornozelo e respostas plantares do extensor (reflexo de Babinski). Usualmente, a função cognitiva é preservada, sendo que o envolvimento dos nervos cranianos não é comum. A apresentação clínica pode se assemelhar a esclerose múltipla ou à mielopatia de infecção por HIV. A situação ideal é que os estudos laboratoriais em pacientes com suspeita de PET/MAH incluam imagens por ressonância magnética (IRM) do cérebro e da medula espinhal, punção lombar e sorologia. Infelizmente, a maioria desses casos ocorre em países com escassez de recursos onde não há disponibilidade de IRM. Foram propostos critérios diagnósticos que classificam a PET/MAH como definitiva, provável ou possível, com fundamento nos sintomas mielopáticos, na sorologia ou na detecção do DNA do HTVL-I e na exclusão de outros distúrbios.71
Observa-se a presença de lesões desmielinizantes na massa branca e nas regiões paraventriculares do cérebro e da medula espinhal. Usualmente, o provírus do HTVL-I não é encontrado nas células do parênquima do SNC, embora possa ser detectado em alguns linfócitos associados ao líquido cerebrospinal. Embora níveis provirais séricos elevados de HTVL-I sejam fatores de risco importantes para o desenvolvimento de mielopatia associada ao HTLV-I (MAH), a presença de provírus no sangue não é suficiente para produzir a doença, sendo que, provavelmente, a imunopatogênese da MAH esteja associada a fatores genéticos de hospedeiro.
Embora o HTLV-I seja reconhecido como a causa principal de leucemia/linfoma de células T do adulto (LTA) e de paraparesia espástica tropical/mielopatia associada ao HTLV-I (PET/MAH), o vírus possivelmente esteja envolvido no desenvolvimento de diversos distúrbios autoimunes diferentes, incluindo artropatia crônica, alveolite pulmonar, síndrome de Sjögren, uveíte, artrite, doença do tecido conjuntivo e polimiosite. Além disso, a doença autoimune da tireoide também foi associada às infecções pelo HTLV-I.
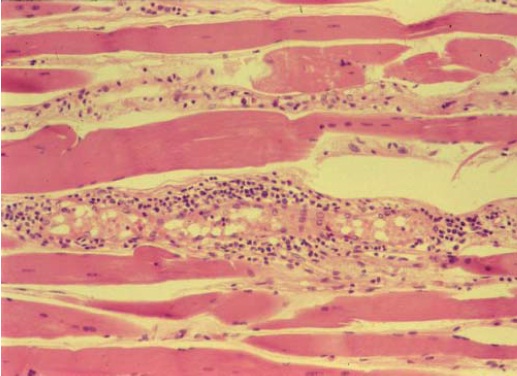
Polimiosite. Existem relatos indicando que há um aumento na prevalência de polimiosite em pacientes infectados pelo HTLV-I.72,73 Os sintomas relatados consistem de febre, astenia, mialgia grave e fraqueza muscular. As descobertas das análises laboratoriais incluem níveis anormais de creatina sérica, níveis anormais de fosfoquinase e do aspartato de aminostransferase, hipergamaglobulinemia policlonal, presença da circulação de complexos imunes e aumento nos níveis dos componentes C3 e C4 do complemento. As alterações miopáticas são evidentes na eletromiografia. As descobertas histopatológicas observadas em espécimes de biópsias musculares incluem infiltração de linfócitos no endomísio e fibras musculares atróficas e necróticas com fagócitos74,75 [ver a Figura 7].
A patogênese da polimiosite em pacientes infectados pelo HTLV-I encontra-se atualmente em fase de investigação. Descobriu-se que as células T CD8+ autorreativas de pacientes positivos para o HTLV-I com polimiosite possuem expansão clonal76 e podem exercer um efeito miotóxico sobre as fibras musculares que expressam moléculas de classe I do complexo de histocompatibilidade principal (MHC, do inglês major histocompatibility complex).77 Os espécimes de biópsia de pacientes com miosite sugerem que a expressão de níveis elevados de autoantígenos e de MHC-I através da regeneração das células musculares resultam na ativação das vias do fator nuclear kB. Essas células musculares podem ser os alvos do ataque por linfócitos T citotóxicos (LTCs).78,79
Figura 7: A amostra de uma biópsia muscular de um paciente com polimiosite grave responsiva a esteroides que foi coinfectado com HIV-1 e com o vírus linfotrópico de células T humanas tipo I (HTLV-I, do inglês human T-cell lymphotropic vírus type I) mostra infiltração linfoide extensiva no músculo. Os produtos genéticos tax/rex do RNA mensageiro do HTLV-I foram detectados no espécime de biópsia.
Uveíte. O HTLV-I pode causar um tipo específico de inflamação intraocular que se caracteriza por reações uveais agudas granulomatosas ou não granulomatosas, opacidades vítreas com irite e vasculite retinal.80,81 Esse tipo de inflamação intraocular pode ser mediada por citocinas produzidas pelas células T CD4+ infectadas pelo HTLV-I.82-84
Na maior parte dos pacientes, as alterações inflamatórias uveais e vasculares retinais respondem aos corticosteroides tópicos ou sistêmicos e desaparecem em algumas semanas.85 As complicações de longo prazo da uveite causada pelo HTLV-I incluem catarata, opacidades vítreas persistentes, degeneração retinocoroidal e neuropatia óptica; provavelmente o prognóstico para a visão não seja muito bom.86-88
Artrite. Há um forte envolvimento do HTLV-I no desenvolvimento de um tipo de artrite que se assemelha à artrite reumatoide (AR) em portadores. Os vírus conhecidos por artritogênicos, como o HTLV-I, produzem artrite inflamatória destrutiva em modelos animais.89 A proteína tax foi identificada como o antígeno mais importante para a proliferação de células sinoviais;90 a prevalência de positividade da proteína tax do HTLV-I em pacientes com artrite reumatoide é três vezes mais elevada do que em doadores de sangue saudáveis.91 Além disso, a expansão oligoclonal de células T foi identificada no sinóvio de pacientes com artrite reumatoide causada pelo HTLV. A expressão do mRNA env e tax do HTLV-I foi detectada no sinóvio afetado e nas células T do sangue periférico de pacientes. A proteína env do HTLV-I também pode desempenhar um papel antigênico importante no desenvolvimento de artropatia em pacientes com artrite crônica associada ao HTLV-I.92
Pneumonite/alveolite linfocítica. Há relatos de pneumonite linfocítica em pacientes com PET/MAH e doenças autoimunes (p.ex., síndrome de Sjögren) e em portadores de HTLV-I sem mielopatias associadas.93,94 Espécimes de biópsias pulmonares de pacientes com HTLV-I mostraram a presença de infiltração linfocítica, aumento na contagem total de células e linfocitose por células T no líquido de lavado broncoalveolar (LBA), fornecendo evidências adicionais de tropismo do HTLV-I para o pulmão.95 O aumento sistêmico e local na carga de DNA proviral do HTLV-I pode estar implicado na patogênese do envolvimento pulmonar em portadores de HTLV-I.94 Há alguns relatos de níveis elevados do receptor solúvel da interleucina 2 (IL-2R) no soro e no BALF em pacientes com PET/MAH causada pelo HTLV-I.96 Levando-se em consideração que as células T do pulmão liberam IL-2R solúvel, levantou-se a hipótese de que os mecanismos imunológicos desempenham um papel importante no desenvolvimento de lesões pulmonares em pacientes portadores do HTLV-I.97,98
Doença tireoidea autoimune. O HTLV-I foi associado ao desenvolvimento de tireoidite de Hashimoto99 e doença de Graves.100 A patogênese e a patobiologia das doenças tireoideas associadas ao HTLV-I ainda não foram estabelecidas, embora a produção de citocinas e de genes celulares por células T infectadas pelo HTLV-I contribua para o desenvolvimento dessas doenças.101
Embora, inicialmente, o HTLV-II tenha sido considerado um patógeno sanguíneo com potencial limitado para a produção de doenças,102,103 estudos subsequentes desafiaram essa visão, principalmente no que diz respeito aos pacientes coinfectados com HTLV-II e HIV.104,105 Assim como o HTLV-I, o HTLV-II causa PET/MAH e doenças neurodegenerativas, porém raramente é encontrado em pacientes com doença linfoproliferativa. A associação entre HTLV-II e malignidade foi observada pela primeira vez em casos de leucemia de células pilosas variantes de células T106 que, subsequentemente, se expandiu através de relatos de casos que descreveram o isolamento do HTLV-II em um paciente com leucemia causada por células T CD8+ e de um paciente portador de HIV que havia sido coinfectado com linfoma cutâneo de células T (CTCL, do inglês cutaneous T cell lymphoma).108
Ainda encontra-se em discussão se o HTLV-II desempenha um papel mais amplo na patogênese de leucemia ou linfoma. Investigações rigorosas excluíram definitivamente o papel do HTLV-II na patogênese da leucemia clássica de células pilosas variantes de células B109,110 e da leucemia granulocítica de células grandes.107
O HTLV-II foi também associado ao desenvolvimento raro de PET/MAH. Em 2004 havia aproximadamente 12 casos conhecidos de mielopatia associada ao HTLV-II,111 incluindo quatro casos de uma coorte prospectiva bem definida de 405 doadores de sangue infectados pelo HTLV-II (prevalência acumulada de 10%; intervalo de confiança de 0,3-2,5) que foram acompanhados por mais de dez anos.112 As evidências epidemiológicas vincularam também a infecção pelo HTLV-II nesses doadores de sangue a um risco elevado de complicações infecciosas, tais como pneumonia, tuberculose e infecções no trato urinário.103
O HTLV-I e o HTLV-II são copatógenos frequentes em indivíduos infectados pelo HIV, principalmente nas grandes áreas metropolitanas onde o uso de drogas injetáveis é um modo comum de transmissão.113,114 Além disso, existem relatos frequentes de coinfecções por HIV-1 e HTLV-I em regiões geográficas de alta soroprevalência de HTLV-I, como o Brasil e a Região do Caribe.115-117 As avaliações clínicas de pacientes coinfectados encontraram uma associação frequente com complicações neurológicas, incluindo mielopatia associada ao HTLV e neuropatia periférica.105,118-120 Os pacientes coinfectados têm também grande probabilidade de apresentar complicações hematológicas, tais como trombocitopenia, assim como infecções virais no sistema respiratório, no trato urinário e hepatite C.119 Embora a terapia antirretroviral altamente ativa (HAART, do inglês highly active antiretroviral therapy) tenha diminuído significativamente a mortalidade associada à AIDS,121 ainda não estão suficientemente claros os benefícios potenciais da HAART em indivíduos infectados pelo HIV.122-124 Um estudo que envolveu 47 pacientes coinfectados pelo HIV e pelo HTLV chegou à conclusão que a HAART não protegeu contra doenças neurológicas e não teve nenhum impacto sobre a carga proviral do HTLV.125 Na realidade, alguns relatórios anedóticos sugerem que a terapia HAART pode aumentar a carga viral do HTLV-I/II.126
Um estudo de coorte de 2004 documentou os resultados clínicos e as probabilidades de sobrevida em pacientes coinfectados pelo HIV-1 e pelo HTLV-I ou pelo HTLV-II.110 Nesse estudo, inexplicavelmente, a coinfecção por HTLV melhorou a sobrevida e retardou a progressão da AIDS. Entretanto, ficou claramente evidente que houve um aumento na frequência de complicações associadas ao HTLV, incluindo PET/MAH e neuropatia periférica.
A dinâmica das coinfecções retrovirais é complexa. Embora o HIV-1, o HTLV-I e o HTLV-II compartilhem um tropismo preferencial por células T, os efeitos virológicos são distintos em cada um desses vírus.127 O HIV-1 é altamente citopático para as células T CD4+, enquanto que HTLV-I e o HTLV-II não são citopáticos e têm um grande potencial para provocar proliferação clonal e transformação de células T.128 Na realidade, a exposição ao HTLV-I ou ao HTLV-II poderá aumentar a infecção produtiva nas células infectadas pelo HIV-1.129,130 No entanto, não há documentação suficiente que comprove o efeito recíproco (i.e., o HIV-1 aumentando o HTLV-I/II).131 Em uma população de pacientes, os níveis de proteínas tax no HTLV-I/II e os níveis de proteínas rex no mRNA eram mais elevados nos pacientes com coinfeção por HIV/HTLV, em comparação com os pacientes infectados apenas por HTLV-I ou HTLV-II.132 As proteínas regulatórias induzidas pelo HIV-1 podem produzir tanto efeitos autócrinos como efeitos parácrinos em microambientes coinfetados pelo HIV-1 e pelo HTLV-I/II, levando a uma suprarregulação na expressão do HTLV-I/II.133,134
Atualmente, o rastreamento de doadores de sangue para verificar a presença de HTLV-I e HTLV-II é um procedimento de rotina em países como Estados Unidos, Canadá, Europa, Japão e em vários países na Região do Caribe.135 O imunoensaio enzimático (EIA, do inglês enzyme immunoassay) é o principal ensaio de rastreamento; o ensaio Western blot é usado como teste de confirmação. Entretanto, o EIA não consegue fazer a distinção entre infecções causadas pelo HTLV-I e pelo HTLV-II por causa da homologia proteica significativa entre os dois vírus. É importante ressaltar que os anticorpos para o HTLV não entram em reação cruzada com as proteínas do HIV. Depois da repetição de resultados positivos com o ensaio EIA, o diagnóstico de infecção por HTLV poderá ser confirmado nas situações em que os anticorpos para os dois produtos genéticos (as proteínas gag e env) forem detectados no ensaio Western blot. Por exemplo, um espécime que demonstrar a reatividade dos anticorpos à p24gag e à pg46env ou à gp61/68env, ou ambas, é considerado positivo para HTLV-I ou HTLV-II.
No caso de espécimes positivos no ensaio EIA, e que reagirem com qualquer uma das bandas do ensaio Western blot, mas não com as proteínas gag e env, os resultados são considerados indeterminados. Os espécimes positivos no ensaio EIA, mas que não apresentarem imunorreatividade a qualquer uma das bandas do ensaio Western blot para o HTLV, são considerados negativos para anticorpos do HTLV-I e do HTLV-II e, nessa hipótese, o resultado do EIA é considerado falso-positivo. Para melhorar a distinção entre HTLV-I e o HTLV-II, o ensaio Western blot foi modificado para incluir proteínas recombinantes específicas para cada tipo a partir de glicoproteínas externas do HTLV-I (rgp46envI) e do HTLV-II (rgp46envII), assim como um peptídeo recombinante truncado – rp21e – da glicoproteína transmembrânica gp21env. Em um dos estudos sobre o tema, os pacientes com perfis indeterminados no ensaio Western blot (i.e., nenhuma reatividade dos anticorpos a p24, p19 ou rp21e), e que não tinham fatores de risco para infecção por HTLV, aparentemente não foram infectados pelo HTLV-I ou pelo HTLV-II pela ampliação da reação em cadeia da polimerase (PCR, do inglês polymerase chain reaction).136 Aparentemente, esses resultados indeterminados do ensaio Western blot representam anticorpos para outros antígenos virais e celulares que entram em reação cruzada com as proteínas do HTVL. Todavia, os doadores de sangue com resultados indeterminados são diferenciados e seu sangue não pode ser usado em transfusões.
A PCR se transformou no método de referência para determinar estados infecciosos, para validar ensaios sorológicos, para fazer a distinção entre HTLV-I e HTLV-II, para estudar in vivo a carga viral e a distribuição tecidual e para fazer avaliações adicionais de pacientes com fatores de risco para infecções causadas pelo HTLV, cujo estado sorológico seja indeterminado ou negativo.135 A PCR é também um método importante para fazer testes de infecção pelo HTLV em lactentes, tendo em vista que seu estado sorológico pode ser indefinido por causa da presença dos anticorpos maternos que foram transferidos passivamente. Além disso, a PCR é importante para a detecção de infecções no período de janela entre infecção e soroconversão.135
Todos os pacientes diagnosticados com leucemia/linfoma de células T de adultos (LTA) devem ser encaminhados para um centro médico terciário especializado no tratamento de malignidades associadas ao HTLV-I. A quimioterapia convencional para tratamento de malignidades linfoides não é eficaz contra formas agressivas de LTA; portanto, o tratamento de LTA se tornou um tema importante para vários estudos clínicos. A combinação de quimioterapias destinadas especificamente para o tratamento de LTA elevou consideravelmente a taxa de respostas de pacientes com LTA, embora não tenha estendido suficientemente a sobrevida mediana observada nos regimes citotóxicos padrões (p.ex., ciclofosfamida, doxorubicina, vincristina e prednisona), que é inferior a dois anos. Um teste atual usado nos Estados Unidos utiliza a administração diária de zidovudina oral em combinação com doses elevadas de interferon alfa.137 O trióxido de arsênio induz a interrupção do ciclo celular e a apoptose das células da LTA in vitro e está sendo avaliado como um agente quimioterápico adjuvante em combinação com o interferon alfa.138 Há diversos relatos no Japão sobre o sucesso do transplante de células-tronco alogênicas após a terapia citorredutiva.139,140 O mogamulizumab, um anticorpo monoclonal humanizado com atividade antitumoral potente, comprovou que é eficiente e bem tolerado em pacientes com LTA recorrente ou refratário. Esse medicamento foi aprovado no Japão para essa indicação e se encontra em fase de testes clínicos nos Estados Unidos e na Europa.141 Os tratamentos adjuvantes para complicações extranodais de LTA incluem o uso de radiação com feixes eternos para tumores líticos ósseos e tumores no interior da medula espinhal; psoraleno mais terapia com raios ultravioleta para lesões causadas por linfoma cutâneo de células T;142 e bifosfonatos para o tratamento de hipercalcemia.
O gerenciamento de doenças infecciosas em pacientes com LTA inclui o uso da profilaxia com trimetoprim-sulfametoxazol contra infecções causadas pelo organismo P. jiroveci em pacientes com a forma aguda da doença e, possivelmente, em pacientes com LTA crônica. Uma das opções aconselhadas é o monitoramento do sangue periférico para verificar o eventual desenvolvimento de antigenemia pelo citomegalovírus (CMV), de modo que a terapia à base de ganciclovir poderá ser iniciada antes do desenvolvimento da pneumonite causada pelo CMV. Os pacientes com LTA com histórico de reatividade à tuberculina, ou que tenham residido em países com taxas elevadas de tuberculose, devem ser considerados para a profilaxia com isoniazida, embora o envolvimento hepático da LTA talvez impeça o uso de medicamentos hepatotóxicos.
Aparentemente, alguns portadores de HTLV-I ou HTLV-II que não tenham LTA também apresentam deficiência imune, evidenciada por uma elevação no risco de complicações de determinadas doenças infecciosas, incluindo estrongiloidíase.143,144 Os indivíduos infectados pelo HTLV-I em áreas altamente endêmicas para o organismo Strongyloides stercoralis provavelmente precisem fazer exame de fezes para verificar a presença de ovos e de parasitas, embora não exista nenhuma recomendação formal a esse respeito. Há também relatos de casos únicos de outras infecções associadas a infecções causadas pelo HTLV-I/HTLV-II, incluindo sarna norueguesa, moluscos contagiosos disseminados e histoplasmose extrapulmonar. Para finalizar, as infecções cutâneas causadas por estafilococos e estreptococos são comuns na síndrome de dermatite infecciosa descrita na Jamaica.145
Os esforços para tratar PET/MAH e HTLV-I associada a doenças autoimunes utilizando-se vários compostos antirretrovirais foram decepcionantes.146 Os corticosteroides e agentes imunossupressivos como a azatioprina possivelmente melhorem a progressão da doença, porém não são adequados para uso no longo prazo por causa dos efeitos adversos.147 Os resultados da imunoterapia com interferon alfa variaram de mínimos a moderados, dependendo do grau da inflamação e do nível de destruição tecidual.148,149
A despeito do desenvolvimento de protótipos de vacinas contra o HTLV-I,150-152 nenhuma das vacinas candidatas chegou a avançar até o nível de testes clínicos. Em parte, talvez esse fato se deva à soroprevalência excessivamente baixa de infecções pelo HTLV-I na maioria das nações prósperas sob o ponto de vista econômico. As vacinas anti- HTLV-II seriam extremamente benéficas em grupos de alto risco como os usuários de drogas injetáveis. O maior benefício de uma vacina não é a proteção contra infecções, mas a ação como um agente imunomodulador no tratamento de LTA latente ou crônica.153
As designações HTLV-III e HTLV-IV foram aplicadas originalmente ao HIV-1 e ao HIV-2, respectivamente; essa nomenclatura foi modificada a partir do momento em que se chegou à conclusão que o HIV-1 e o HIV-2 são lentivírus, ao passo que os HTLVs são oncornavírus.154-156 Atualmente, os oncornavírus conhecidos como HTLV-III e HTLV-IV foram identificados entre caçadores na África Central.3,157,158 Provavelmente esses vírus tenham resultado do contato direto de caçadores-coletores com o sangue de primatas não humanos, tendo em vista que, sob o ponto de vista genético, esses vírus são muito semelhantes aos vírus linfotrópicos de células T de primatas que foram isolados de um grande variedade de primatas não humanos nas áreas rurais da República dos Camarões.158,159 Embora, atualmente, o HTLV-III e o HTLV-IV estejam restritos a uma pequena população em uma área remota, o deslocamento de populações, como resultado do desmatamento e da migração da população rural para áreas urbanas, poderá promover a disseminação desses vírus. Até o presente momento, não se conhece o potencial exato para doenças causadas por esses retrovírus humanos recentemente descobertos.160
Os vírus tipo espuma, também conhecidos por vírus espumosos, se caracterizam pela capacidade de induzir efeitos citopáticos típicos em células cultivadas, com formação de sincícios.161 Os vírus espumosos possuem transcriptase reversa e compartilham homologia genética com outros vírus da família Retroviridae.162 Aparentemente, esses vírus infectam uma ampla variedade de animais, incluindo felinos e primatas não humanos;163 sabe-se que quase todas as espécies de primatas não humanos são infectadas pelos vírus espumosos dos símios.164
Em 1971 foi documentado o relato do HFV isolado de um espécime de biópsia de carcinoma nasofaríngeo,165 sendo que o trabalhos subsequentes demonstraram que, sob o ponto de vista genético, esse vírus foi intimamente relacionado a um vírus espumoso de chimpanzés.166 Estudos sorológicos realizados na Europa e na África indicam que há uma prevalência muito baixa de sororreatividade aos vírus espumosos em seres humanos.162,167 Assim como ocorre com o HTLV-III e o HTLV-IV, a transmissão de infecções causadas pelo vírus espumoso humano podem ser consequência do contato com tecidos e líquidos corporais de primatas não humanos.168 Até o presente momento ainda não foi identificada nenhuma ligação causativa entre o vírus espumoso e doenças em seres humanos.
Os retrovírus endógenos humanos (RVEHs) se localizam no interior do genoma humano, porém, de maneira geral, não existem como provírus completamente organizados. Porções das sequências retrovirais, como gag, pol, e env (assim como outras sequências genéticas estruturais e regulatórias, conhecidas como retroelementos), são incorporadas sob a forma de um complexo no interior do DNA eucariótico.169 Consequentemente, os RVEHs são replicados juntamente com os genes celulares de hospedeiro. Dependendo da inteireza dos retroelementos do RVEH, possivelmente sejam expressas partículas retrovirais defeituosas ou competentes com replicação completa. Por exemplo, os provírus do RVEH contendo os genes gag e promotor, porém sem um gene pol ou env efetivo, têm um grande potencial para adquirir a capacidade para construir uma partícula semelhante a um vírus que não poderia germinar a partir da membrana celular, tendo em vista que não possuem invólucro. Existem poucos RVEHs que contêm quase todos os genes virais, como os membros da família RVEH-K.170 Os provírus defeituosos, expressos em alguma células teratogênicas, expressam diversas proteínas, incluindo gag, cORF, protease, polimerase e env, e podem se juntar formando partículas virais.171 Os membros da família RVEH-K se transcrevem sob a regulação das respectivas repetições terminais longas e expressam RNA viral em níveis baixos em vários tecidos e tumores humanos.171 Consequentemente, postula-se que desempenhem algum papel na patogênese desses tumores.
A expressão das proteínas do RVEH podem induzir autoimunidade através de diversos mecanismos.172 Por exemplo, as proteínas do RVEH possivelmente simulem antígenos de hospedeiro ou ajam como superantígenos, resultando na ativação e na proliferação de células T. A expansão clonal de populações de células T poderia produzir citocinas (incluindo TNF-a, IL-1a e IL-1ß), conforme foi demonstrado em alguns estudos, para suprarregular a expressão do mRNA do retrovírus endógeno humano.173
As partículas retrovirais humanas foram observadas no interior de amostras de glândulas salivares coletadas em pacientes portadores da síndrome de Sjögren e de alguns pacientes soronegativos para o HIV com deficiência de células T CD4+.174,175 Alguns estudos de soroprevalência sugeriram que a reatividade dos anticorpos a esses RVEHs está presente em um percentual elevado de pacientes portadores de doenças autoimunes, como o lúpus eritematoso sistêmico.176 Ainda não está suficientemente claro até que ponto esses vírus ou suas subunidades contribuem para a patogênese da doença.172
Mark A. Beilke, MD e Christy Barrios, PhD e John Heinegg não mantêm nenhuma relação comercial com os fabricantes dos produtos ou com os fornecedores de serviços mencionados neste capítulo. Daniel R. Kuritzkes, MD, FACP atuou como consultor da Bristol Myers-Squibb, Celera, Gilead, GlaxoSmithKline, Human Genome Sciences, InnaVirVax, Koronis Pharma, Inc, Merck Sharp & Dolme Corp, subsidiária da Merck & Co.,Inc, Tobira Therapeutics e ViiV Healthcare; e recebeu apoio financeiro para as pesquisas da Gilead e da Merck Sharp & Dolme Corp, subsidiária da Merck & Co.,Inc.
1. Baltimore D. Reversal of information flow in the growth of RNA tumor viruses. N Engl J Med 1971;284:273.
2. Kalyanaraman VS, Sarngadharan MG, Robert-Guroff M, et al. A new subtype of human T-cell leukemia virus (HTLV-II) associated with a T-cell variant of hairy cell leukemia. Science 1982;218:571.
3. Mahieux R, Gessain A. New human retroviruses: HTLV-3 and HTLV-4. Med Trop 2005;65:525.
4. Calattini S, Chevalier SA, Duprez R, et al. Discovery of a new human T-cell lymphotropic virus (HTLV-3) in Central Africa. Retrovirology 2005;2:30.
5. Levy JA. Endogenous C-type viruses: double agents in natural life processes. Biomedicine 1976;24:84.
6. Abraham GN, Khan AS. Human endogenous retroviruses and immune disease. Clin Immunol Immunopathol 1990;56:1.
7. Aaronson SA, Dunn CY. Endogenous C-type viruses of BALB/c cells: frequencies of spontaneous and chemical induction. J Virol 1974;13:181.
8. Meiering CD, Linial ML. Historical perspective of foamy virus epidemiology and infection. Clin Microbiol Rev 2001;14:165.
9. Heneine W, Schweizer M, Sandstrom P, et al. Human infection with foamy viruses. Curr Top Microimmunol 2003;277:181.
10. Linial M. Why aren’t foamy viruses pathogenic? Trends Microbiol 2000;8:284.
11. Smith DK, Neal JJ, Holmberg SD. Unexplained opportunistic infections and CD4+ T-lymphocytopenia without HIV infection. N Engl J Med 1993;328:373.
12. Ho DD, Cao Y, Zhu T, et al. Idiopathic CD4+ T-lymphocytopenia immunodeficiency without evidence of HIV infection. N Engl J Med 1993;328:380.
13. Gupta S, Ribak CE, Gollapudi S, et al. Detection of a human intracisternal retroviral particle associated with CD4+ T-cell deficiency. Proc Natl Acad Sci U S A 1992;89:7831.
14. Liu HF, Goubau P, Van Brussel M, et al. The three human T-lymphotropic virus type I subtypes arose from three geographically distinct simian reservoirs. J Gen Virol 1996;77:359.
15. Gao F, Bailes E, Robertson DL, et al. Origin of HIV-1 in the chimpanzee Pan troglodytes troglodytes. Nature 1999;397:436.
16. Kaplan JE, Khabbaz RF. The epidemiology of human T-lymphotropic virus type I and II. Rev Med Virol 1993;3:137.
17. Dezzutti CS, Lal RB. Human T-cell lymphotropic virus types I and II. In: Murray PR, Baron KJ, Pfaller MA, et al, editors. Manual of clinical microbiology. Washington (DC): American Society for Microbiology; 1999. p. 871.
18. Schreiber GB, Murphy EL, Horton JA, et al. Risk factors for human T-cell lymphotropic virus types I and II (HTLV-I and -II) in blood donors. NHLBI Retrovirus Epidemiology Donor Study. J Acquir Immune Defic Syndr Hum Retrovirol 1997;14:263.
19. Schreiber GB, Busch MP, Kleinman SH, et al. The risk of transfusion-transmitted viral infections. The Retrovirus Epidemiology Donor Study. N Engl J Med 1996;334:1685.
20. Green PL, Chen SY. Human T-cell leukemia virus types 1 and 2. In: Knipe DM, Howley PM, editors. Fields virology. Vol 2. Philadelphia: Lippincott Williams & Wilkins; 2001. p. 1941.
21. Goff SP. Retroviridae: the retroviruses and their replication. In: Knipe DM, Howley MD, editors. Fields virology. Vol 2. Philadelphia: Lippincott Williams & Wilkins; 2001. p. 1871.
22. Dalton AJ. RNA tumor viruses: terminology and ultrastructural aspects of virion morphology and replication. J Natl Cancer Inst 1972;49:323.
23. Bouillant AM, Becker SA. Ultrastructural comparison of Oncovirinae (type C), Spumavirinae, and Lentivirinae: three subfamilies of Retrovirridae found in farm animals. J Natl Cancer Inst 1984;72:1075.
24. Fine D, Schochetman G. Type D primate retroviruses: a review. Cancer Res 1978;38:3123.
25. Lo RC, Howatson AF. Identification of type C viruses by electron microscopy. Cancer Res 1978;38:932.
26. de Giuli C, Hanafusa H, Kawai S, et al. Relationship between A-type and C-type particles in cells infected by Rous sarcoma virus. Proc Natl Acad Sci U S A 1975;72:3706.
27. Palmer E, Goldsmith CS. Ultrastructure of human retroviruses. J Electron Microsc Tech 1988;8:3.
28. Levy JA, Hoffman AD, Kramer SM, et al. Isolation of lymphocytopathic retroviruses from San Francisco patients with AIDS. Science 1984;225:840.
29. Orenstein JM. Ultrastructural pathology of human immunodeficiency virus infection. Ultrastruct Pathol 1992;16:179.
30. Arthur LO, Bess JW Jr, Sowder RC 2nd, et al. Cellular proteins bound to immunodeficiency viruses: implications for pathogenesis and vaccines. Science 1992;258:1935.
31. Dezzutti CS, Lal RB. Human T-cell lymphotropic virus types I and II. In: Murray PR, Baron KJ, Pfaller MA, et al, editors. Manual of clinical microbiology. Washington (DC): American Society for Microbiology; 1999. p. 871.
32. Vidal AU, Gessain A, Yoshida M, et al. Phylogenetic classification of human T cell leukaemia/lymphoma virus type I genotypes in five major molecular and geographical subtypes. J Gen Virol 1994;75:3655.
33. Eiraku N, Novoa P, da Costa Ferreira M, et al. Identification and characterization of a new and distinct molecular subtype of human T-cell lymphotropic virus type 2. J Virol 1996;70:1481.
34. Goon PK, Igakura T, Hanon E, et al. Human T cell lymphotropic virus type 1-specific CD4+ T cells: immunodominance hierarchy and preferential infection with HTLV-I. J Immunol 2004;172:1735.
35. Ijichi S, Ramundo MB, Takahashi H, et al. In vivo cellular tropism of human T cell leukemia virus type II (HTLV-II). J Exp Med 1992;176:293.
36. Lal RB, Owen SM, Rudolph DL, et al. In vivo cellular tropism of human T-lymphotropic virus type II is not restricted to CD8+ cells. Virology 1995;210:441.
37. Jassal SR, Pohler RG, Brighty DW. Human T-cell leukemia virus type 1 receptor expression among syncytium-resistant cell lines revealed by a novel surface glycoprotein-immunoadhesin. J Virol 2001;75:8317.
38. Trejo SR, Ratner L. The HTLV receptor is a widely expressed protein. Virology 2000;268:41.
39. Manel N, Kim FJ, Kinet S, et al. The ubiquitous glucose transporter GLUT-1 is a receptor for HTLV. Cell 2003;115:449.
40. Jones KS, Fugo K, Petrow-Sadowski C, et al. Human T-cell leukemia virus type 1 (HTLV-I) and HTLV-II use different receptor complexes to enter T cells. J Virol 2006;80:8291.
41. Bangham CR. The immune control and cell-to-cell spread of human T-lymphotropic virus type 1. J Gen Virol 2003;84:3177.
42. Igakura T, Stinchcombe JC, Goon PK, et al. Spread of HTLV-I between lymphocytes by virus-induced polarization of the cytoskeleton. Science 2003;299:1713.
43. Ohshima K, Ohgami A, Matsuoka M, et al. Random integration of HTLV-I provirus: increasing chromosomal instability. Cancer Lett 1998;132:203.
44. Luciw PA. Human immunodeficiency viruses and their replication. In: Fields BN, Knipe DM, Howley PM, editors. Fields virology. Vol 2. Philadelphia: Lippincott-Raven; 1996. p. 1881.
45. Flint SJ, Enquist LW, Krug RM, et al. Principles of virology: molecular biology, pathogenesis, and control. Washington (DC): American Society of Microbiology; 2000.
46. Bangham CR. The immune control and cell-to-cell spread of human T-lymphotropic virus type 1. J Gen Virol 2003;84:3177.
47. Oyayama A, Chen YM, Tachibana N, et al. High incidence of antibodies to HTLV-I tax in blood relatives of adult T cell leukemia virus patients. J Infect Dis 1991;163:47.
48. Manns A, Blattner WA. The epidemiology of the human T-cell lymphotrophic virus type I and type II: etiologic role in human disease. Transfusion 1991;31:67.
49. Blanchard S, Astier-Gin T, Moynet D, et al. Different HTLV-I neutralization patterns among sera of patients infected with cosmopolitan HTLV-I. Virology 1998;245:90.
50. Astier-Gin T, Portail JP, Londos-Gagliardi D, et al. Neutralizing activity and antibody reactivity toward immunogenic regions of the human T cell leukemia virus type I surface glycoprotein in sera of infected patients with different clinical states. J Infect Dis 1997;175:716.
51. Sundaram R, Lynch MP, Rawale SV, et al. De novo design of peptide immunogens that mimic the coiled coil region of human T-cell leukemia virus type-1 glycoprotein 21 transmembrane subunit for induction of native protein reactive neutralizing antibodies. J Biol Chem 2004;279:2414.
52. de Thé G, Kazanji M. An HTLV-I/II vaccine: from animal model to clinical trials? J Acquir Immune Defic Syndr Hum Retrovirol 1996;13:191.
53. Takamoto T, Makino M, Azuma M, et al. HTLV-I–infected T cells activate autologous CD4+ T cells susceptible to HTLV-I infection in a costimulatory molecule-dependent fashion. Eur J Immunol 1997;27:1427.
54. Hanon E, Hall S, Taylor GP, et al. Abundant tax protein expression in CD4+ T cells infected with human T-cell lymphotropic virus type I (HTLV-I) is prevented by cytotoxic T lymphocytes. Blood 2000;95:1386.
55. Kubota R, Kawanishi T, Matsubara H, et al. Demonstration of human T lymphotropic virus type I (HTLV-I) tax-specific CD8+ lymphocytes directly in peripheral blood of HTLV-I–associated myelopathy/tropical spastic paraparesis patients by intracellular cytokine detection. J Immunol 1998;161:482.
56. Ureta-Vidal A, Pique C, Garcia Z, et al. Human T cell leukemia virus type I (HTLV-I) infection induces greater expansions of CD8 T lymphocytes in persons with HTLV-I–associated myelopathy/tropical spastic paraparesis than in asymptomatic carriers. J Infect Dis 2001;183:857.
57. Bieganowska K, Hollsberg P, Buckle GJ, et al. Direct analysis of viral specific CD8+ T cells with soluble HLA-A2/Tax11-19 tetramer complexes in patients with human T cell lymphotropic virus-associated myelopathy. J Immunol 1999;162:1765.
58. Yashiki S, Fujiyoshi T, Arima N, et al. HLA-A*26, HLA-B*4002, HLA-B*4006, and HLA-B*4801 alleles predispose to adult T cell leukemia: the limited recognition of HTLV type 1 tax peptide anchor motifs and epitopes to generate anti-HTLV type 1 tax CD8(+) cytotoxic T lymphocytes. AIDS Res Hum Retroviruses 2001;17:1047.
59. Arnulf B, Thorel M, Poirot Y, et al. Loss of the ex vivo but not the reinducible CD8+ T-cell response to Tax in human T-cell leukemia virus type 1-infected patients with adult T-cell leukemia/lymphoma. Leukemia 2004;18:126.
60. Nomura M, Ohashi T, Nishikawa K, et al. Repression of tax expression is associated both with resistance of human T-cell leukemia virus type 1–infected T cells to killing by tax-specific cytotoxic T lymphocytes and with impaired tumorigenicity in a rat model. J Virol 2004;78:3827.
61. Furukawa Y, Kubota R, Tara M, et al. Existence of escape mutant in HTLV-I tax during the development of adult T-cell leukemia. Blood 2001;97:987.
62. Ohashi T, Hanabuchi S, Kato H, et al. Prevention of adult T-cell leukemia–like lymphoproliferative disease in rats by adoptively transferred T cells from a donor immunized with human T-cell leukemia virus type 1 Tax-coding DNA vaccine. J Virol 2000;74:9610.
63. Ahmed R, Morrison LA, Knipe DM. Persistence of viruses. In: Fields BN, Knipe DM, Howley PM, editors. Fields virology. Vol 2. Philadelphia: Lippincott-Raven; 1996. p. 219.
64. Shimoyama M. Diagnostic criteria and classification of clinical subtypes of adult T-cell leukaemia-lymphoma: a report from the Lymphoma Study Group (1984–87). Br J Haematol 1991;79:428.
65. Blattner W, Manhattan C. Human T-cell lymphotropic virus types I and II. In: Mandell GL, Bennett JE, Dolin R, editors. Mandell, Douglas, and Bennett’s principles and practice of infectious diseases. 6th ed. Philadelphia: Churchill Livingstone; 2005. p. 2098.
66. Fouchard N, Flageul B, Bagot M, et al. Lack of evidence of HTLV-I/II infection in T CD8 malignant or reactive lymphoproliferative disorders in France: a serological and/or molecular study of 169 cases. Leukemia 1995;9:2087.
67. Wood GS, Schaffer JM, Boni R, et al. No evidence of HTLV-I proviral integration in lymphoproliferative disorders associated with cutaneous T-cell lymphoma. Am J Pathol 1997;150:667.
68. Roman GC, Roman LN. Tropical spastic paraparesis: a clinical study of 50 patients from Tumaco (Colombia) and review of the worldwide features of the syndrome. J Neurol Sci 1988;87:121.
69. Cleghorn FR, Blattner WA. Human T-cell lymphotropic virus and HTLV infection. In: Holmes KK, Sparling PF, Mardh PA, et al, editors. Sexually transmitted diseases. New York: McGraw-Hill; 1999. p. 259.
70. Iwasaki Y. Pathology of chronic myelopathy associated with HTLV-I infection (HAM/TSP). J Neurol Sci 1990;96:103.
71. De Castro-Costa CM, Araujo AQ, Barreto MM, et al. Proposal for diagnostic criteria of tropical spastic paraparesis/HTLV-I–associated myelopathy (TSP/HAM). AIDS Res Hum Retroviruses 2006;22:931.
72. Mora CA, Garruto RM, Brown P, et al. Seroprevalence of antibodies to HTLV-I in patients with chronic neurological disorders other than tropical spastic paraparesis. Ann Neurol 1988;23 Suppl:192.
73. Sowa JM. Human T lymphotropic virus I, myelopathy, polymyositis and synovitis: an expanding rheumatic spectrum. J Rheumatol 1992;19:316.
74. Khanlou H, Mechain C, Maunand B, et al. Polymyositis in HTLV-I infected patient. J Intern Med 1996;239:371.
75. Inose M, Higuchi I, Yoshimine K, et al. Pathological changes in skeletal muscle in HTLV-1–associated myelopathy. J Neurol Sci 1992;110:73.
76. Saito M, Higuchi I, Saito A, et al. Molecular analysis of T cell clonotypes in muscle-infiltrating lymphocytes from patients with human T lymphotropic virus type 1 polymyositis. J Infect Dis 2002;186:1231.
77. Dalakas MC. Mechanisms of disease: signaling pathways and immunobiology of inflammatory myopathies. Nat Clin Pract Rheumatol 2006;2:219.
78. Nagaraju K, Casciola-Rosen L, Lundberg I, et al. Activation of the endoplasmic reticulum stress response in autoimmune myositis: potential role in muscle fiber damage and dysfunction. Arthritis Rheum 2005;52:1824.
79. Nagaraju K. Role of major histocompatibility complex class I molecules in autoimmune myositis. Curr Opin Rheumatol 2005;17:725.
80. Mochizuki M, Ono A, Ikeda E, et al. HTLV-I uveitis. J Acquir Immune Defic Syndr Hum Retrovirol 1996;13 Suppl 1:S50.
81. Beilke MA. Human T-cell lymphotropic virus type I–associated uveitis in an African American. Am J Ophthalmol 1995;119:104.
82. Yoshimura K, Mochizuki M, Araki S, et al. Clinical and immunologic features of human T-cell lymphotropic virus type I uveitis. Am J Ophthalmol 1993;116:156.
83. Sagawa K, Mochizuki M, Masuoka K, et al. Immunopathological mechanisms of human T cell lymphotropic virus type 1 (HTLV-I) uveitis: detection of HTLV-I-infected T cells in the eye and their constitutive cytokine production. J Clin Invest 1995;95:852.
84. Ono A, Mochizuki M, Yamaguchi K, et al. Immunologic and virologic characterization of the primary infiltrating cells in the aqueous humor of human T-cell leukemia virus type–1 uveitis: accumulation of the human T-cell leukemia virus type–1–infected cells and constitutive expression of viral and interleukin-6 messenger ribonucleic acids. Invest Ophthalmol Vis Sci 1997;38:676.
85. Nakao K, Ohba N. Clinical features of HTLV-I associated uveitis. Br J Ophthalmol 1993;77:274.
86. Nakao K, Ohba N, Nakagawa M, et al. Clinical course of HTLV-I–associated uveitis. Jpn J Ophthalmol 1999;43:404.
87. Takahashi T, Takase H, Urano T, et al. Clinical features of human T-lymphotropic virus type 1 uveitis: a long-term follow-up. Ocul Immunol Inflamm 2000;8:235.
88. Yukawa E, Urano T, Nakahara M, et al. Pattern-reversal visual evoked potentials in patients with human T-lymphotropic virus type 1 uveitis. Curr Eye Res 2006;31:37.
89. Masuko-Hongo K, Kato T, Nishioka K. Virus-associated arthritis. Best Pract Res Clin Rheumatol 2003;17:309.
90. Hasunuma T, Sumida T, Nishioka K. Human T cell leukemia virus type–I and rheumatoid arthritis. Int Rev Immunol 1998;17:291.
91. Zucker-Franklin D, Pancake BA, Brown WH. Prevalence of HTLV-I Tax in a subset of patients with rheumatoid arthritis. Clin Exp Rheumatol 2002;20:161.
92. Kato T, Asahara H, Kurokawa MS, et al. HTLV-I env protein acts as a major antigen in patients with HTLV-I–associated arthropathy. Clin Rheumatol 2004;23:400.
93. Kompoliti A, Gage B, Sharma L, et al. Human T-cell lymphotropic virus type 1–associated myelopathy, Sjögren syndrome, and lymphocytic pneumonitis. Arch Neurol 1996;53:940.
94. Mori S, Mizoguchi A, Kawabata M, et al. Bronchoalveolar lymphocytosis correlates with human T lymphotropic virus type I (HTLV-I) proviral DNA load in HTLV-1 carriers. Thorax 2005;60:138.
95. Sugimoto M, Mita S, Tokunaga M, et al. Pulmonary involvement in human T-cell lymphotropic virus type–I uveitis: T-lymphocytosis and high proviral DNA load in bronchoalveolar lavage fluid. Eur Respir J 1993;6:938.
96. Sugimoto M, Nakashima H, Matsumoto M, et al. Pulmonary involvement in patients with HTLV-I–associated myelopathy: increased soluble IL-2 receptors in bronchoalveolar lavage fluid. Am Rev Respir Dis 1989;139:1329.
97. Kimura I. HABA (HTLV-I associated bronchiolo-alveolar disorder). Jpn J Thorac Dis 1992;30:787.
98. Kawai H, Kashiwagi S, Sano Y, et al. HTLV-I associated myelopathy with Hashimoto’s thyroiditis: a report of two cases. Rinsho Shinkeigaku 1991;31:648.
99. Kawai H, Inui T, Kashiwagi S, et al. HTLV-I infection in patients with autoimmune thyroiditis (Hashimoto’s thyroiditis). J Med Virol 1992;38:138.
100. Sarui H, Sano A, Maruyama T, et al. Severe Graves ophthalmopathy accompanied by HTLV-I–associated uveitis (HAU) and anti-neutrophil cytoplasmic antibody-related vasculitis: a case report and a review of the literature of HAU with Graves disease. Am J Med Sci 2002;324:109.
101. Matsuda T, Tomita M, Uchihara JN, et al. Human T cell leukemia virus type-I–infected patients with Hashimoto’s thyroiditis and Graves’ disease. J Clin Endocrinol Metab 2005;90:5704.
102. Manns A, Blattner WA. The epidemiology of the human T-cell lymphotrophic virus type I and type II: etiologic role in human disease. Transfusion 2001;31:67.
103. Murphy EL, Glynn SA, Fridey J, et al. Increased incidence of infectious diseases during prospective follow-up of human T-lymphotropic virus type II- and I-infected blood donors. Retrovirus Epidemiology Donor Study. Arch Intern Med 1999;159:1485.
104. Berger JR, Svenningsson A, Raffanti S, et al. Tropical spastic paraparesis–like illness occurring in a patient dually infected with HIV-1 and HTLV-II. Neurology 1991;41:85.
105. Zehender G, De Maddalena C, Osio M, et al. High prevalence of human T cell lymphotropic virus type II infection in patients affected by human immunodeficiency virus type 1–associated predominantly sensory polyneuropathy. J Infect Dis 1995;172:1595.
106. Kalyanaraman VS, Sarngadharan MG, Robert-Guroff M, et al. A new subtype of human T-cell leukemia virus (HTLV-II) associated with a T-cell variant of hairy cell leukemia. Science 1982;218:571.
107. Loughran TP Jr, Coyle T, Sherman MP, et al. Detection of human T-cell leukemia/lymphoma virus, type II, in a patient with large granular lymphocyte leukemia. Blood 1992;80:1116.
108. Poiesz B, Dube D, Dube S, et al. HTLV-II–associated cutaneous T-cell lymphoma in a patient with HIV-1 infection. N Engl J Med 2000;342:930.
109. Loughran TP Jr, Sherman MP, Ruscetti FW, et al. Prototypical HTLV-I/II infection is rare in LGL leukemia. Leuk Res 1994;18:423.
110. Rosenblatt JD, Giorgi JV, Golde DW, et al. Integrated human T-cell leukemia virus II genome in CD8+ T cells from a patient with “atypical” hairy cell leukemia: evidence for distinct T and B cell lymphoproliferative disorders. Blood 1988;71:363.
111. Araujo A, Hall WW. Human T-lymphotropic virus type II and neurological disease. Ann Neurol 2004;56:10.
112. Orland JR, Engstrom J, Fridey J, et al. Prevalence and clinical features of HTLV neurologic disease in the HTLV outcomes study. Neurology 2003;61:1588.
113. Khabbaz RF, Onorato IM, Cannon RO, et al. Seroprevalence of HTLV-I and HTLV-II among intravenous drug users and persons in clinics for sexually transmitted diseases. N Engl J Med 1992;326:375.
114. Ciancianaini P, Magnani G, Barchi E, et al. Serological and clinical follow-up of Italian IVDU cohort of HTLV-II/HIV-1 co-infected patients: evidence of direct relationship between CD4 count and HTLV-II proviral load [abstract]. AIDS Res Human Retrovirus 2001;17 Suppl:152.
115. Carneiro-Proietti AB, Lima-Martins MV, Passos VM, et al. Presence of human immunodeficiency virus (HIV) and T-lymphotropic virus type I and II (HTLV-I/II) in a haemophiliac population in Belo Horizonte, Brazil, and correlation with additional serological results. Haemophilia 1998;4:47.
116. Bartholomew C, Blattner W, Cleghorn F. Progression to AIDS in homosexual men co-infected with HIV and HTLV-I in Trinidad. Lancet 1987;2:1469.
117. Figueroa JP, Ward E, Morris J, et al. Incidence of HIV and HTLV-1 infection among sexually transmitted disease clinic attenders in Jamaica. J Acquir Immune Defic Syndr Hum Retrovirol 1997;15:232.
118. Beilke MA, Greenspan DL, Impey A, et al. Laboratory study of HIV-1 and HTLV-I/II coinfection. J Med Virol 1994;44:132.
119. Beilke MA, Theall KP, O’Brien M, et al. Clinical outcomes and disease progression among patients coinfected with HIV and human T lymphotropic virus types 1 and 2. Clin Infect Dis 2004;39:256.
120. Harrison LH, Vaz B, Taveira DM, et al. Myelopathy among Brazilians co-infected with HTLV-I and HIV. Neurology 1997;48:13.
121. Murphy EL, Collier AC, Kalish LA, et al. Highly active antiretroviral therapy decreases mortality and morbidity in patients with advanced HIV disease. Viral Activation Transfusion Study Investigators. Ann Intern Med 2001;135:17.
122. Garcia-Lerma JG, Nidtha S, Heneine W. Susceptibility of human T cell leukemia virus type 1 to reverse-transcriptase inhibitors: evidence for resistance to lamivudine. J Infect Dis 2001;184:507.
123. Hill SA, Lloyd PA, McDonald S, et al. Susceptibility of human T cell leukemia virus to nucleoside reverse transcriptase inhibitors. J Infect Dis 2003;188:424.
124. Machuca A, Soriano V. In vivo fluctuation of HTLV-I and HTLV-II proviral load in patients receiving antiretroviral drugs. J Acquir Immune Defic Syndr 2000;24:189.
125. Silva MT, Neves ES, Grinsztejn B, et al. Neurological manifestations of coinfection with HIV and human T-lymphotropic virus type 1. AIDS 2012;26:521.
126. Turci M, Pilotti E, Ronzi P, et al. Coinfection with HIV-1 and HTLV-II in intravenous drug users is associated with delayed progression to AIDS. J Acquir Immune Defic Syndr 2006;41:100.
127. Gallo RC. Human retroviruses after 20 years: a perspective from the past and prospects for their future control. Immunol Rev 2002;185:236.
128. Blattner W. Human T lymphotrophic viruses: HTLV-I and HTLV-II. In: Broder S, Merrigan TC, Bolognesi D, editors. Textbook of AIDS medicine. Philadelphia: Lippincott Williams & Wilkins; 1994. p. 887.
129. Cheng H, Tarnok J, Parks WP. Human immunodeficiency virus type 1 genome activation induced by human T-cell leukemia virus type 1 Tax protein is through cooperation of NF-kappaB and Tat. J Virol 1998;72:6911.
130. Moriuchi H, Moriuchi M, Fauci AS. Factors secreted by human T lymphotropic virus type I (HTLV-I)–infected cells can enhance or inhibit replication of HIV-1 in HTLV-I–uninfected cells: implications for in vivo coinfection with HTLV-I and HIV-1. J Exp Med 1998;187:1689.
131. De Rossi A, Saggioro D, Calabro ML, et al. Reciprocal activation of human T-lymphotropic viruses in HTLV-I–transformed cells superinfected with HIV-1. J Acquir Immune Defic Syndr 1991;4:380.
132. Beilke MA, Japa S, Vinson DG. HTLV-I and HTLV-II virus expression increases with HIV-1 coinfection. J Acquir Immune Def Syndr 1997;17:391.
133. Ensoli B, Buanaguro L, Barillari G, et al. Release, uptake and effects of extracellular human immunodeficiency virus type 1 Tat protein on cell growth and viral transactivation. J Virol 1993;67:277.
134. Kubota S, Furuta RA, Hatanaka M, et al. Modulation of HTLV-I gene expression by HIV-1 Rev through an alternative RxRE-independent pathway mediated by the RU5 portion of the 5’-LTR. Biochem Biophys Res Commun 1998;243:79.
135. Dezzutti CS, Lal RB. Human T-cell lymphotropic virus types I and II. In: Murray PR, Baron KJ, Pfaller MA, et al, editors. Manual of clinical microbiology. Washington (DC): American Society for Microbiology; 1999. p. 871.
136. Lal RB, Rudolph DL, Coligan JE, et al. Failure to detect evidence of human T-lymphotropic virus (HTLV) type I and type II in blood donors with isolated gag antibodies to HTLV-I/II. Blood 1992;80:544.
137. Ramos JC, Ghosh SK, Toomey LL, et al. Successful treatment with AZT and IFN in adult T-cell leukemia-lymphoma is characterized by in vivo induction of TRAIL and AP-1 activity. AIDS Res Hum Retroviruses 2003;19 Suppl:26.
138. Nasr R, Rosenwald A, El-Sabban ME, et al. Arsenic/interferon specifically reverses 2 distinct gene networks critical for the survival of HTLV-I–infected leukemic cells. Blood 2003;101:4576.
139. Abe Y, Yashiki S, Choi I, et al. Eradication of virus-infected T-cells in a case of adult T-cell leukemia/lymphoma by nonmyeloablative peripheral blood stem cell transplantation with conditioning consisting of low-dose total body irradiation and pentostatin. Int J Hematol 2002;76:91.
140. Ishikawa T. Current status of therapeutic approaches to adult T-cell leukemia. Int J Hematol 2003;78:304.
141. Subramaniam JM, Whiteside G, McKeage K, et al. Mogamulizumab: first global approval. Drugs 2012;72:1293.
142. Kaplan MH, Hall WW, Susin M, et al. Syndrome of severe skin disease, eosinophilia, and dermatopathic lymphadenopathy in patients with HTLV-II complicating human immunodeficiency virus infection. Am J Med 1991;91:300.
143. Marsh BJ. Infectious complications of human T cell leukemia/lymphoma virus type I infection. Clin Infect Dis 1996;23:138.
144. Satoh M, Toma H, Sugahara K, et al. Involvement of IL-2/IL-2R system activation by parasite antigen in polyclonal expansion of CD4(+)25(+) HTLV-I–infected T-cells in human carriers of both HTLV-I and S. stercoralis. Oncogene 2002;21:2466.
145. La Grenade L, Schwartz RA, Janniger CK. Childhood dermatitis in the tropics: with special emphasis on infective dermatitis, a marker for infection with human T-cell leukemia virus-I. Cutis 1996;58:115.
146. Taylor GP, Goon PK, Usuku K, et al. The Bridge Study: a double-blind placebo controlled trial of zidovudine plus lamivudine for treatment of patients with HTLV-I associated myelopathy. AIDS Res Hum Retroviruses 2003;19 Suppl:13.
147. Nakagawa M, Nakahara K, Maruyama Y, et al. Therapeutic trials in 200 patients with HTLV-I–associated myelopathy/tropical spastic paraparesis. J Neurovirol 1996;2:345.
148. Feng J, Misu T, Fujihara K, et al. Interferon-alpha significantly reduces cerebrospinal fluid CD4 cell subsets in HAM/TSP. J Neuroimmunol 2003;141:170.
149. Saito M, Nakagawa M, Kaseda S, et al. Decreased human T lymphotropic virus type I (HTLV-I) provirus load and alteration in T cell phenotype after interferon-alpha therapy for HTLV-I–associated myelopathy/tropical spastic paraparesis. J Infect Dis 2004;189:29.
150. Sundaram R, Lynch MP, Rawale SV, et al. De novo design of peptide immunogens that mimic the coiled coil region of human T-cell leukemia virus type-1 glycoprotein 21 transmembrane subunit for induction of native protein reactive neutralizing antibodies. J Biol Chem 2004;279:24141.
151. Sundaram R, Sun Y, Walker CM, et al. A novel multivalent human CTL peptide construct elicits robust cellular immune responses in HLA-A*0201 transgenic mice: implications for HTLV-I vaccine design. Vaccine 2003;21:2767.
152. Kazanji M, Tartaglia J, Franchini G, et al. Immunogenicity and protective efficacy of recombinant human T-cell leukemia/lymphoma virus type 1 NYVAC and naked DNA vaccine candidates in squirrel monkeys (Saimiri sciureus). J Virol 2001;75:5939.
153. Kannagi M, Harashima N, Kurihara K, et al. Adult T-cell leukemia: future prophylaxis and immunotherapy. Expert Rev Anticancer Ther 2004;4:369.
154. Popovic M, Sarngadharan MG, Read E, et al. Detection, isolation and continuous production of cytopathic retroviruses (HTLV-III) from patients with AIDS and pre-AIDS. Science 1984;224:497.
155. Kanki PJ, Hopper JR, Essex M. The origins of HIV-1 and HTLV-4/HIV-2. Ann N Y Acad Sci 1987;511:370.
156. Hahn BH, Kong LI, Lee SW, et al. Relation of HTLV-4 to simian and human immunodeficiency viruses. Nature 1987;300:184.
157. Calattini S, Chevalier SA, Duprez R, et al. Discovery of a new human T-cell lymphotropic virus (HTLV-3) in Central Africa. Retrovirology 2005;2:30.
158. Wolfe ND, Heneine W, Carr JK, et al. Emergence of unique primate T-lymphotropic viruses among central African bushmeat hunters. Proc Natl Acad Sci U S A 2005;102:7994.
159. Switzer WM, Qari SH, Wolfe ND, et al. Ancient origin and molecular features of the novel human T-lymphotropic virus type 3 revealed by complete genome analysis. J Virol 2006;80:7427.
160. Gessain A, Rua R, Betsem E, et al. HTLV-3/4 and simian foamy retroviruses in humans: discovery, epidemiology, cross-species transmission and molecular virology. Virology 2013;435:187.
161. Aguzzi A. The foamy virus family: molecular biology, epidemiology and neuropathology. Biochim Biophys Acta 1993;1155:1.
162. Meiering CD, Linial ML. Historical perspective of foamy virus epidemiology and infection. Clin Microbiol Rev 2001;14:165.
163. Hooks JJ, Gibbs CJ Jr. The foamy viruses. Bacteriol Rev 1975;39:169.
164. Heneine W, Schweizer M, Sandstrom P, et al. Human infection with human foamy viruses. Curr Top Microimmunol 2003;277:81.
165. Achong BF, Mansell PW, Epstein MA, et al. An unusual virus in cultures from a human nasopharyngeal carcinoma. J Natl Cancer Inst 1971;46:299.
166. Brown P, Nemo G, Gajdusek DC. Human foamy virus: further characterization, seroepidemiology, and relationship to chimpanzee foamy viruses. J Infect Dis 1978;137:421.
167. Schweizer M, Turek R, Hahn A, et al. Markers of foamy virus infections in monkeys, apes, and accidentally infected humans: appropriate testing fails to confirm suspected foamy virus prevalence in humans. AIDS Res Hum Retrovirus 1995;11:161.
168. Goepfert PA, Ritter G Jr, Peng X, et al. Analysis of West African hunters for foamy virus infections. AIDS Res Hum Retrovirus 1996;12:1725.
169. Löwer R, Löwer J, Kurth R. The viruses in all of us: characteristics and biological significance of human endogenous retrovirus sequences. Proc Natl Acad Sci U S A 1996;93:5177.
170. Ono M, Yasunaga T, Miyata T, et al. Nucleotide sequence of human endogenous retrovirus genome related to the mouse mammary tumor virus genome. J Virol 1986;60:589.
171. Tönjes RR, Löwer R, Boller K, et al. HERV-K: the biologically most active human endogenous retrovirus family. J Acquir Immune Defic Syndr 1996;13 Suppl:261.
172. Portis JL. Minireview: perspectives on the role of endogenous human retroviruses in autoimmune diseases. Virology 2002;296:1.
173. Katsumata K, Ideda H, Sato M, et al. Cytokine regulation of env gene expression of human endogenous retrovirus-R in human vascular endothelial cells. Clin Immunol 1999;93:75.
174. Garry RF, Fermin CD, Hart DJ, et al. Detection of human intracisternal A-type retroviral particle antigenically related to HIV. Science 1990;250:1127.
175. Gupta S, Ribak CE, Gollapudi S, et al. Detection of a human intracisternal retroviral particle associated with CD4+ T-cell deficiency. Proc Natl Acad Sci U S A 1992;89:7831.
176. Sander DM, Szabo S, Gallaher WR, et al. Involvement of human intracisternal A-type retroviral particles in autoimmunity. Microsc Res Tech 2005;68:222.
177. Biggar RJ, Buskell-Bales Z, Yakshe PN, et al. Antibody to human retroviruses among drug users in three east coast American cities. J Infect Dis 1991;163:57.
178. Briggs NC, Battjes RJ, Cantor KP, et al. Seroprevalence of HTLV-2 infection, with or without HIV-1 coinfection, among US intravenous drug users. J Infect Dis 1995;172:51.
179. Hall WW, Ishak R, Zhu SW, et al. Human T lymphotropic virus type II (HTLV-II): epidemiology, molecular properties, and clinical features of infection. J Acquir Immune Defic Syndr Hum Retrovirol 1996;13 Suppl:204.
180. Salemi M, Lewis M, Egan JF, et al. Different population dynamics of human T cell lymphotropic virus type II in intravenous drug users compared with endemically infected tribes. Proc Natl Acad Sci U S A 1999;96:13253.
O MedicinaNET é o maior portal médico em português. Reúne recursos indispensáveis e conteúdos de ponta contextualizados à realidade brasileira, sendo a melhor ferramenta de consulta para tomada de decisões rápidas e eficazes.
| Medicinanet Informações de Medicina S/A Cnpj: 11.012.848/0001-57 | info@medicinanet.com.br |
MedicinaNET - Todos os direitos reservados.
Em função da pandemia do Coronavírus informamos que não estaremos prestando atendimento telefônico temporariamente. Permanecemos com suporte aos nossos inscritos através do e-mail info@medicinanet.com.br.